PART ONE—
HUMANS AND ANIMALS
One—
Introduction
James J. Sheehan
In 1810, William Blake painted a picture that came to be known as Adam Naming the Beasts . Blake's portrait of the first man reminds us of a Byzantine icon of Christ: calm, massive, and immobile, Adam dominates the science. One of his arms is raised in an ancient gesture signifying speech, while around the other a serpent meekly coils. In the background, animals move in an orderly, peaceful file. Of course, we know the harmony depicted here will not last. Soon Adam and his progeny will lose their serence place in nature; no longer will they be comfortable in their sovereignty over animals or secure in the unquestioned power of their speech. Our knowledge of what is coming gives blake's picture is special, melancholy power.[1]
Since the Fall, man's place in nature has always been problematic. The problems begin with Genesis itself, where the story of creation is told twice. In the second chapter, the source of Blake's picture, God creates Adam and then all other living things, which are presented to man "to see what he would call them: and whatsoever Adam called every living creature, that was the name thereof," The first chapter, however, has a somewhat different version of these events. Here man is the last rather than the first creature to be made; while still superior to the rest by his special relationship to God, man nevertheless appears as part of a larger natural order. A similar ambiguity can be found in Chapter Nine, which begins with a divine promise to Noah that all beings will fear him but then goes on to describe a covenant between God and Noah and "every living creature of all flesh." From the very start of the Judeo-Christian tradition, therefore, humanity is at once set apart from, and joined with, the realm of other living things.[2]
In Greek cosmology, humanity's relationship to animals was yet more
uncertain. Like the Hebrews, the Greeks seemed eager to establish human hegemony over nature. Animals sacrifice, which was so central to Greek religion, ritually affirmed the distinctions between humans and beasts, just as it sought to establish connections between the human and divine. Aristotle, the first great biologist to speculate about human nature, developed an elaborate hierarchy of living beings, in which all creatures—beginning with human females—were defined on a sliding scale that began with adult males. But the line between humanity and its biological neighbors was more permeable for Greeks than for Hebrews. Gods frequently took on animals form, which allowed them to move about the world in disguise. As punishment, humans could be turned into beasts. Moreover, a figure like Heracles, who was human but with supernatural connections, expressed his association with animals by wearing skins on his body and a lion's head above his own. And if animal sacrifice set humans and animals apart, there were other rituals that seemed to blur the distinction between them. Dionysus's Maenads, for instance, lived wild and free, consumed raw flesh, and knew no sexual restraint. By becoming like beasts, the Maenads achieved a "divine delirium" and thus direct contact with the gods.[3]
Christians saw nothing godlike in acting like a beast. To them, the devil often appeared in animal form, a diabolic beast or, as Arnold Davidson points out, in some terrible mix of species. Bestiality, sexual transgression across the species barrier, was officially regarded as the worst sin against nature; it remained a capital crime in England until the second half of the nineteenth century. Humanity's proper relationship to animals was that of master; beasts existed to serve human needs. "Since beasts lack reason," Saint Augustine taught, "we need not concern ourselves with their sufferings," an opinion echoed by an Anglican bishop in the seventeenth century who declared, "We may put them [animals] to any kind of death that the necessity either of our food or physic will require." Even those who took a softer view of humanity's relationship with animals believed that our hegemony over the world reflected our special ties to the creator. "Man not only rules the animals by force," the Renaissance philosopher, Ficino, wrote, "he also governs, keeps and teaches them. Universal providence belongs to God, who is the universal cause. Hence man who generally provides for all things, both living and lifeless, is a kind of God."[4]
Although set apart from the rest of creation by their privileged relationship with God, many Christians felt a special kinship to animals. As Keith Thomas shows in his splendid study, Man and the Natural Workd, so close were the ties of people to the animals among whom they lived that often "domestic beasts were subsidiary members of the human community." No less important than these pressing sympathies of everyday inter-
dependence were the weight of cultural habit and the persistent power of half-forgotten beliefs. Until well into the eighteenth century, many Furopeans viewed the world anthropomorphically, imposing on animals human traits and emotions, holding them responsible for their "crimes," admiring them for their alleged expressions of pious sentiment. Although condemned by the orthodox and ridiculed by secular intellectuals, belief in the spirituality of animals persisted. As late as the 1770s, an English clergyman could write, "I firmly believe that beasts have souls; souls truly and properly so-called."[5]
By the end of the eighteenth century, such convictions were surely exceptional among educated men and women. The expansion of scientific knowledge since the Renaissance had helped to produce a view of the world in which there seemed to be little room for animal souls. The great classification schemes of the late seventeenth and eighteenth centuries encouraged rational, secular, and scientific conceptions of the natural order. As a result, the anthropomorphic attitudes that had invested animals—and even plants—with human characteristics gradually receded; nature was now seen as something apart from human affairs, a realm to be studied and mastered with the instruments of science. Here is Thomas's concise summary of this process:
In place of a natural world redolent with human analogy and symbolic meaning, and sensitive to man's behavior, they [the natural scientists] constructed a detached natural scene to be viewed and studied by the observer from the outside, as if by peering through a window, in the secure knowledge that the objects of contemplation inhabited a separate realm, offering no omens or signs, without human meaning or significance.[6]
Within this new world, humans' claims to hegemony were based on their own rational faculties rather than divine dispensation. Reason became the justification as well as the means of humanity's mastery. Because they lack reason, Descartes argued, animals were like machines, without souls, intelligence, or feeling. Animals do not act independently, "it is nature that acts in them according to the arrangement of their organs, just as we see how a clock, composed merely of wheels and springs, can reckon the hours." Rousseau agreed. Every animal, he wrote in A Discourse on Inequality, was "only an ingenious machine to which nature has given sense in order to keep itself in motion and protect itself." Humans are not in thrall to their instincts and senses; unlike beasts, when nature commands, humans need not obey. Free will, intellect, and above all, the command of language gives people the ability to choose, create, and communicate.[7]
Not every eighteenth-century thinker was sure that humanity's unquestioned uniqueness had survived the secularization of the natural
order. Lord Bolingbroke, for example, still regarded man as "the principal inhabitant of this planet" but cautioned against making too much of humanity's special status.
Man is connected by his nature, and therefore, by the design of the Author of all Nature, with the whole tribe of animals, and so closely with some of them, that the distance between his intellectual faculties and theirs, which constitutes as really, though not so sensibly as figure, the difference of species, appears, in many instances, small, and would probably appear still less, if we had the means of knowing their motives, as we have of observing their actions.
When Alexander Pope put these ideas into verse, he added that the sin of pride that brought Adam's fall came not from his approaching too close to God but rather from drawing too far away from other living things:
Pride then was not, nor arts that pride to aid;
Man walk'd with beast, joint tenants of the shade;
And it was Pope who best expressed the lingering anxiety that must attend humanity's position between gods and beasts:
Plac'd in this isthmus of a middle state,
A being darkly wise and rudely great,
With too much knowledge for the sceptic side,
With too much weakness for the stoic pride,
He hangs between; in doubt to act or rest;
In doubt to deem himself a good or beast;
In doubt his Mind or Body to prefer;
Born but to die, and reas'ning but or err. . . .
Chaos of Thought and Passion all confus'd,
Still by himself abus'd, or disabus'd;
Created half to rise, and half to fall,
Great lord of all things, yet a prey to all;
Sole judge of Truth, in endless error hurl'd;
The glory, jest, and riddle of the world.[8]
In the second half of the eighteenth century, what Pope had once called the "vast chain of being" began to be seen as dynamic rather than static, organic rather than mechanistic. The natural order now seemed to be the product of a sosmic evolution that, as Kant wrote in 1755, "is never finished or complete." This meant that everything in the universe—from species of plants and animals to the structure of distant galaxies and the earth's surface—was produced by and subject to powerful forces of change. By the early nineteenth century, scientists had
begun to examine this evolutonary process in a systematic fashion. In 1809, Jean Baptiste de Lamarck published Philosophie zoologique, which set forth a complex theory to explain the transformation of species over time. Charles Lyell, whose Principles of Geology began to appear in 1830, doubted biological evolution but offered a compelling account of the earth's changing character through the long corridors of geologic time. Thus was the stage set for the arrival of Darwin's Origin of Species, by far the most famous and influential of all renditions of "temporalized chains of being." With this book, Darwin provides the basis for our view of the natural order and thus links eighteenth-century cosmology and twentieth-century evolutionary biology.[9]
Even before Darwin formulated his theory of natural selection, he seems to have sensed this his scientific observations would have powerful implications for the relationship between humans and animals. In a notebook entry of 1837, Darwin first sketched what would beocme a central theme in his life's work:
If we choose to let conjecture run wild, then animals, our fellow brethren in pain, diseases, death, suffering and famine—our slaves in the most laborious works, our companions in our amusements—they may partake [of] our origins in one common ancestor—we may be all netted together.
When he published Origin of Species twenty-two years later, he approached the matter delicately and indirectly: "it does not seem incredible" that animals and plants developed from lower forms, "and if we admit this, we must likewise admit that all the organic beings which have ever lived on this earth may be descended from some one primordial form." In any event, he promised that in future research inspired by the theory of natural selection, "much light will be thrown on the origin of man and his history."[10]
The meaning of Darwinism for human history quickly moved to the center of the controversies that followed the publication of Origin . In 1863, T. H. Huxley, for example, produced Man's Place in Nature . Darwin himself turned to the evoluton of humanity in The Descent of Man, where he set out to demonstrate that "there is no fundamental difference between man and the higher mammals in their mental faculties." Each of those characteristics once thought to be uniquely human turn out to be shared by higher animals, albeit in lesser degrees: animals cannot speak, but they do communicate; they are intellectually inferior to humans, but they "possess some power of reasoning"; they cannot work as we do, but they can even use tools in a rudimentary way. Darwin's discussion of animal self-consciousness is worth quoting at length, not simply because it conveys the flavor of his argument but also because it illustrates the
ease with which he slipped from talking about differences between humans and animals to describing differences between "races" of men.
It may be freely admitted that no animal is self-conscious, if by this term it is implied that he reflects on such points, as whence he comes or whither he will go, or what is life and death, and so forth. But how can we feel sure that an old dog with excellent memory and some power of imagination, as shewn by his dreams, never reflects on his past pleasures and pains in the chase? And this would be a form of self-consciousness. On the other hand, as Büchner has remarked, how little can the hard-worked wife of a degraded Australian savage, who uses very few abstract words, and cannot count above four, exert her self-consciousness, or reflect on the nature of her own existence.
Darwin carried on his examination of humans and animals in The Expression of the Emotions in Man and Animals. Here he seeks to demonstrate that basic emotions have common origins and a biological base by pointing out the similarity of emotional expressions among human societies and between humans and animals. Like reason, language, and consciousness, emotions such as love, fear, and shame are not the sole and undisputed property of humanity.[11]
The research program suggested in this work on human and animal emotions was not immediately taken up by Darwin's many disciples. There were, to be sure, many who sought to apply Darwinism to human society, but they usually did so without a systematic examination of the resemblances between human and animal behavior. To social scientists influenced by Darwin, what mattered was less that people were animals than that they were still—in Harriet Ritvo's phrase—"the top animals," separated from the rest by what Dawwin himself had called man's "noble qualities" and "godlike intellect." Most of the natural scientists who followed Darwin turned in the opposite direction, away from humans toward other species with longer evolutionary histories and more accessible biological structures. As a result, empirical work on the connection between human and animal behavior, so central to Darwin's work on emotions, did not become an important part of his legacy until the second half of the twentieth century.[12]
The direct heirs of Darwin's research on human and animal emotion were scientists who studied behavioral biology, the discipline that would come to be called ethology. Although important research on ethology had been conducted during the 1920s and 1930s, the subject did not become prominent until the 1960s; in 1973, three leading ethologists shared the Nobel Prize. The new interest in ethology encouraged the publication of a variety of works of quite uneven quality, but all shared the conviction that studying the similarity of behavior existing across
animal species could yield important insights into the character of human individuals and groups. For example, Konrad Lorenz, a pioneer in the field and one of the Nobel laureates, maintained that, in both man and animals, aggressive behavior was instinctual. "Like the triumph ceremony of the greylag goose, militant enthusiasm in man is a true autonomous instinct: It has its own releasing mechanisms, and like the sexual urge or any other strong instinct, it engenders a specific feeling of intense satisfaction." Such deeply rooted instincts, ethnologists warned, will inevitably pose problems for, and set limits on, humans' ability to control themselves and their societies.[13]
By the 1970s, popular and scientific interest in ethnology had become enmeshed with a larger and more ambitious set of ideas and research enterprises conventinally called sociobiology. Edward O. Wilson, perhaps sociobiology's most vigorous exponent, describes it as the "scientific study of the biological basis of all forms of social behavior in all kinds of organisms, including man." Wilson began to define his view of the field in the concluding chapter of The Insect Societies (1971), which called for the application of his work on the population biology and zoology of insects to vertebrate animals. Four years later, he concluded Sociobiology: The New Synthesis with a chapter entitled "Man: From Sociobiology to sociology." This was followed in 1978 with the more popularly written Oh Human Nature, which examined what Wilson regarded as "four of the elemental categories of behavior, aggression, sex, altruism, and religion," from a sociobiological perspective. Wilson brought an ethnologist's broad knowledge of animal behavior to these subjects but added his own growing concern for the genetic basis of instincts and adaptations. This genetic dimension has become increasingly important in Wilson's most recent work.[14]
Sociobiology in general and Wilson in particular have been the subject of intense attacks from a variety of directions. In the course of these controversies, the term has tended to become a catchall for a variety of different developments in behavioral biology. Like many other controversial movements, sociobiology often seems more solid and coherent to its opponents, who can easily define what they oppose, than to its advocates, who have some trouble agreeing on what they have in common. It is worth noting, for example, that Melvin Konner, while sympathetic to Wilson in many ways, explicitly denies tht his own work, The Tangled Wing (1982), is sociobiology. But what Konner and the sociobiologists do share is the belief that most studies of human beings have been too "anthropocentric." If, as Wilson and others claim, "homo sapiens is a conventional animal species," then there is much to be learned by viewing the human experience as part of a broader biological continuum. Doing so will help us to understand what Darwin, in the dark passage at
the end of The Descent of Man, referred to as "the indelible stamp of his lowly origin" which man still carries in his body and what Konner, in a contemporary version of the same argument, calls the "biological constraints on the human spirit."[15]
We will return to some of the questions raised by sociobiology in the conclusion to this volume. For the moment, it is enough to point out that the conflicts surrounding it—illustrated by the works of Konner and Dupré in the following section—are new versions of ancient controversies. These controversies, while informed by our expandng knowledge of the natural world and expressed in the idiom of our scientific culture, have at thier core our persistent need to define what it means to be human, a need that leads us, just as it led the authors of Genexis, to confront our kinship with and differences from animals.
Two—
The Horror of Monsters*
Arnold I. Davidson
As late as 1941, Lucien Febvre, the great French historian, could complain that there was no history of love, pity, cruelty, or joy. He called for "a vast collective investigation to be opened on the fundamental sentiments of man and the forms they take."[1] Although Febvre did not explicitly invoke horror among the sentiments to be investigated, a history of horror can, as I hope to show, function as an irreducible resource in uncovering our forms of subjectivity.[2] Moreover, when horror is coupled to monsters, we have the opportunity to study systems of thought that are concerned with the relation between the orders of morality and of nature. I will concentrate here on those monsters that seem to call into question, to problematize, the boundary between humans and other animals. In some historical periods, it was precisely this boundary that, under certain specific conditions that I shall describe, operated as one major locus of the experience of horror. Our horror at certain kinds of monsters reflects back to us a horror at, or of, humanity, so that our horror of monsters can provide both a history of human will and subjectivity and ahistory of scientific classifications.
The history of horror, like the history of other emotions, raises extraordinarily difficult philosophical issues. When Febvre's call was answered, mainly by his French colleagues who practiced the so-called history of mentalities, historians quickly recognized that a host of historiographical and methodological problems would have to be faced. No one has faced these problems more directly, and with more profound results, than Jean Delumeau in his monumental two-volume history of fear.[3] But these are issues to which we must continually return. What will be required to write the history of an emotion, a form of sensibility, or type of affectivity, Any such history would require an investigation of
gestures, images, attitudes, beliefs, language, values, and concepts. Furthermore, the problem quickly arose as to how one should understand the relationship between elite and popular culture, how, for example, the concepts and language of an elite would come to be appropriated and transformed by a collective mentality.[4] This problem is especially acute for the horror of monsters, since so many of the concepts I discuss which are ncessary to out understanding of monsters come from high culture—scientific, philosophical, and theological texts. To what extent is the experience of horror, when expressed in a collective mentality, given from by these concepts? Without even attempting to answer these questions here, I want to insist that a history of horror, at both the level of elite concepts and collective mentality, must emphasize the fundamental role of description. We must describe, in much more detail than is usually done, the concepts, attitudes, and values required by and manifested in the reaction of horror. And it is not enough to describe these components piecemeal; we must attempt to retrieve their coherence, to situate them in the structures of which they are a part.[5] At the level of concepts, this demand requires that we reconstruct the rules that govern the relationships between concepts; thus, we will be able to discern the highly structured, rule-governed conceptual spaces that are overlooked if concepts are examined only one at a time.[6] At the level of mentality, we are required to place each attitude, belief, and emotion in the context of the specific collective consciousness of which it forms part.[7] At both levels, we will have to go beyond what is said or expressed in order to recover the conceptual spaces and mental equipment without which the historical texts will lose their real significance.
In 1523, Martin Luther and Phillip Melancthon published a pamphlet entitled Deuttung der czwo grewlichen Figuren, Bapstesels czu Rom und Munchkalbs czu Freyerbeg ijnn Meysszen funden .[8] It was enormously influential and was translated into French, with Calvin's endorsement, in 1557, and into English in 1579 under the title Of two wonderful popish monsters . The pamphlet consisted of a detailed interpretation of two monsters: a pope-ass, discussed mainly by Melancthon, supposedly left on the banks of the Tiber River in 1496, and a monk-calf, interpreted by Luther, that was born on December 8, 1522, in Freiburg (figs. 2.1, 2.2). Both of these monsters were interpreted within the context of a polemic against the Roman church. They were prodigies, signs of God's wrath against the Church which prophesied its imminent ruin. There were two dimensions to the Lutheran exegesis of these monsters.[9] On the one hand, there is a prophetic or eschatological dimension, only diffidently mentioned in this pamphlet, in which monsters and prodigies, as a general phenomenon, were taken to be signs of fundamental changes about to

Fig. 2.1.
The Pope-Ass.
affect the world. Often these signs were interpreted as nothing less than an announcement that the end of the world was at hand, and support for this prophetic interpretation was adduced by citing the Book of Daniel, a biblical text invokec by both Melancthon and Luther. The other dimension, which, following Jean Céard, we can call allegorical, is the one with which this pamphlet is most preoccupied. The allegorical exegesis of these monsters is intended to show that each monster has a very specific

Fig. 2.2.
The Monk-Calf.
interpretation that can be grasped because, in one way or another, it is represented before our eyes in the constitution of the monster itself; each monster is a divine hieroglyphic, exhibiting a particular feature of God's wrath. So, for instance, the pope-ass, according to Melancthon, is the image of the Church of Rome; and just as it is awful that a human body should have the head of an ass, so it is likewise horrible that the Bishop of Rome should be the head of the Church. Similarly, the overly
large ears of the calf-monk exhibit God's denouncement of the practice of hearing confessions, so important to the monks, while the hanging tongue shows that their doctrine is nothing but frivolous prattle.
A useful study could be made of the adjectives that appear in this text; in lieu of such a study, let me just note that "horrible" and "abominable" occur frequently in both Luther's and Melancthon's discussions, often modifying "monster." The mood of these adjectives is accurately conveyed in the translator's introduction to the 1579 English translation of the text. It begins:
Among all the things that are to be seen under the heavens (good Christian reader) there is nothing can stir up the mind of man, and which can engender more fear unto the creatures than the horrible monsters, which are brought forth daily contrary to the works of Nature. The which the most times do note and demonstrate unto us the ire and wrath of God against us for our sins and wickedness, that we have and do daily commit against him.[10]
John Brooke goes on to tell us that his motive for translating this pamphlet is the better "to move the hearts of every good Christian to fear and tremble at the sight of such prodigious monsters,"[11] and he warns his readers not to interpret these two monsters as if they were but fables. He closes his preface with the hope that, after reading this pamphlet, we shall "repent in time from the bottom of our hearts of our sins, and desire him [God] to be merciful unto us, and ever to keep and defend us from such horrible monsters."[12] He concludes with a few more specific remarks about the pope-ass and calf-monk addressed, and we shall not overlook the form of the address, "unto all which fear the Lord."
In order to better understand the preoccupation and fascination with monsters during the sixteenth century, a fascination fastened onto by Luther and Melancthon, whose text is fully representative of an entire genre, we must place these discussions within a wider context. As Jean Delumeau has argued in the second volume of his history of fear, it is within "the framework of a global pessimistic judgment on a time of extreme wickedness that one must place the copious literature dedicated to monsters and prodigies between the end of the fifteenth century and the beginning of the seventeenth."[13] Sinfulness was so great that the sins of men extended to nature itself which, with God's permission and for the instruction of sinners, seemed to have been seized by a strange madness; the resulting monsters were to be understood as illustrations of these sins. Heresy and monsters were frequently linked during this period, both by reformers and Catholics alike. Prodigies were not only specific punishments for particular sins but they also announced greater punishments to come—war, famine, and perhaps even the end of the
world. This proliferation of monsters presaged a dark future explained by God's wrath at the increase of wickedness on earth.[14] Français Belleforest summarized the shared sensibility: "The present time is more monstrous than natural."[15]
To make as clear as possible the relationship between horror and monsters, I am going to focus primarily on one text, Ambroise Paré's Des monstres et prodiges, originally published in 1573 and frequently reprinted after that.[16] Since I am going to set the conceptual context for my discussion of Paré in a rather unconventional way, I want to state explicitly that a full comprehension of this treatise requires that it be placed in relation to other learned and popular treatises on monsters that both preceded and followed it. We are fortunate in this respect to have Céard's thorough treatment of Paré in his brilliant La Nature et les prodiges and in the notes to his critical edition of Des monstres et prodiges;[17] moreover, in the best English-language treatment of monsters, Katharine Park and Lorraine Daston have provided a three-stage periodization—monsters as divine prodigies, as natural wonders, and as medical examples for comparative anatomy and embryology—that is indispensable in helping us to understand shifts in the conceptualization and treatment of monsters from the Middle Ages to the eighteenth century.[18] Rather than summarizing the work of these scholars, I am going to turn to a different kind of text to prepare my discussion of Paré, namely, Thomas Aquinas's Summa Theologica .
Aquinas's Summa is not only the greatest work of genius produced by medieval moral theology but also a profound synthesis of previous work, coherently connecting doctrines, ideas, and arguments whose relationships had never been made very clear; moreover, the Summa also made conceptually determinate notions that had had a deep and wide-ranging significance in the Middle Ages but that had not really been approached with sufficient analytical precision. I am going to use one portion of the Summa as representative of medieval attitudes, attitudes that have lasted, in one form or another, for many subsequent centuries. I shall not address the question of the Summa 's originality in this area; suffice it to say that I believe that this is one place in which Aquinas gave a conceptually powerful formulation to a set of dieas that had been essential, even if not very precise, to most of medieval moral theology.
Part II of Part II, Questions 13 and 154, of the Summa Theologica deal with lust and the parts of lust, respectively. Aquinas begins, in Article 2 of Question 154, by considering the question of whether the venereal act can be without sin. He argues as follows: if the dictate of reason makes use of certain things in a fitting manner and order for the end to which they are adapted, and if this end is truly good, then the use of these things in such a fitting manner and order will not be a sin. Now the
preservation of the bodily nature of an individual is a true good, and the use of food is directed to the preservation of life in the individual. Similarly, the preservation of the nature of the human species is a very great good, and the use of the venereal act is directed to the preservation of the whole human race. Therefore, Aquinas concludes,
wherefore just as the use of food can be without sin, if it be taken in due manner and order, as required for the welfare of the body, so also the use of venereal acts can be without sin, provided they be performed in due manner and order, in keeping with the end of human procreation.[19]
He proceeds in the first article of Question 154 to differentiate six species of lust—simple fornication, adultery, incest, seduction, rape, and the vice contrary to nature—all of which are discussed in the remaining articles.
My concern is with the vices contrary to nature, which are discussed in Articles 11 and 12. In Article 11, he argues that this type of vice is a distinct species of lust, since it involves a special kind of deformity; vices contrary to nature are not only contrary to right reason, as are all the lustful vices, but are also "contrary to the natural order of the venereal act as becoming to the human race," which order has as its end the generation of children.[20] Aquinas distinguishes four categories of vice contrary to nature—bestiality, sodomy, which he interprets as male/male or female/female copulation, the sin of self-abuse, and not observing the natural manner of copulation. It is difficult to determine exactly what falls under this last category, but it is clear from II-II, Question 154, Article 12, Reply to Objection 4, that male/female anal and oral copulation are two of the most grievous ways of not observing the right manner of copulation.
In Article 12, Aquinas rank-orders, from worst to least worst, all of the lustful vices. He claims, first, that all four categories of vice contrary to nature are worse than any of the other vices of lust. So that bestiality, sodomy, not observing the natural manner of copulation, and self-abuse are worse, because of their special deformity, than adultery, rape of a virgin, incest, and so on.[21] Vices contrary to nature are worse in kind and not merely in degree than other lustful vices. Aquinas then goes on to rank-order the vices contrary to nature. The least bad of these vices is self-abuse, since the "gravity of a sin depends more on the abuse of a thing than on the omission of the right use."[22] Next worse is the sin of not observing the right manner of copulation, and this sin is more grievous if the abuse concerns the right vessel than if it affects the manner of copulation in respect of other circumstances. Third worse is sodomy, since use of the right sex is not observed. Finally, the most grievous of all the vices contrary to nature, and so the most grievous of any lustful vice,
is bestiality, since the use of the due species is not observed; moreover, in this instance, Aquinas explicitly cites a biblical text as support.[23] One final remark of Aquinas's must be mentioned before I turn to Paré. About the vices contrary to nature, from masturbation to bestiality, Aquinas writes,
just as the ordering of right reason proceeds from ne, so the order of nature is from God Himself: wherefore in sins contrary to nature, whereby the very order of nature is violated, an injury is done to God, the Author of nature.[24]
To act contrary to nature is nothing less than to act directly contrary to the will of God.
One may understandably be wondering how this discussion of Aquinas is relevant to the treatment of monsters, so let me turn immediately to Paré's Des monstres et prodiges . The preface to his book begins as follows:
Monsters are things that appear outside the course of Nature (and are usually signs of some forthcoming misfortune), such as a child who is born with one arm, another who will have two heads, and additional members over and above the ordinary.
Prodigies are things which happen that are completely against Nature, as when a woman will given birth to a serpent, or to a dog, or some other thing that is totally against Nature, as we shall show hereafter through several examples of said monsters and prodigies.[25]
Cérd has argued that Paré was somewhat indifference to the problem of precisely how one should distinguish monsters from prodigies. Monsters and prodigies did not constitute absolutely separate classes and during the successive editions of his book, Cérd things Paré became more and more convinced that the term "monster" was sufficient to designate all of these phenomena.[26] But however imprecise and unarticulated this distinction might appear, the idea that there was a separate class of phenomena, prodigies, that were completely against nature affected the language, attitude, and conceptualization with which Paré approached his examples.
In the first chapter of Des monstres et prodiges , Paré distinguishes thirteen causes of monsters, which causes, although not completely exhaustive, are all the ones he is able to adduce with assurance. Ten of these causes are straightforwardly natural causes; two, the glory of God and the wrath of God, are straightforwardly supernatural causes; and one, demons and devils, has a long and complicated classificatory history.[27] Briefly, to classify the products of demons and devils as a result of supernatural causes was to threaten to place the devil on a par with God, granting him the same powers to overturn the natural order that God
possessed. The possibility of such a theologically untenable position led to detailed discussions concerning the status of demonic causation; and as we can see from chapter 26 to 34, Paré fit squarely into these discussions, concerned both to grant the reality of the devil and yet to limit his powers. Of the two straightforwardly supernatural causes, Paré's threatment of the first, the glory of God, is exhausted by one example, the restoration of a blind man's sight by Jesus Christ, an example literally copied from Pierre Boaistuau's Histoires Prodigieuses , first published in 1560.[28]
The other supernatural cause, the wrath of God, is far more interesting for my purposes; most of the example produced by Paŕ to illustrate this category are of the same kind, and they are closely linked to the natural cause of the mixture or mingling of seed. I want to discuss these examples in detail in order to support some claims about the history of horror. But I should make one more preliminary remark. Paré, like virtually every writer during this period, had no intellectual difficulty in referring to both supernatural and natural causes; he felt no incompatibility in discussing these two types of cause together. Yet although God was always in the background of Des monstres et prodiges , by far the most space is devoted to natural causes, with God's explicit appearances being relatively few. This contrasts, for instance, with Jacob Rueff's De conceptu et generatione hominis , a book known to Paré, published in 1554 and for a long time the classic work on the problems of generation. Rueff also discussed supernatural and natural causes together, but in Book V of De conceptu , when he discusses monstrous births, Rueff considers them all as divine punishment, and their physical causes, however active, are almost ignored in favor of the evidence of the judgments of God. In Rueff's text, whether the physical or natural causes of the production of monsters, monsters are first of all punishments inflicted by God on sinners.[29] So Paré's book already demonstrates a shift of emphasis that makes his treatment of supernatural causes all the more interesting.
Paré's chapter on thw wrath of God opens with these words:
There are other creatures which astonish us doubly because they do not proceed from the above mentioned causes, but from a fusing together of strange species, which render the creature not only monstrous but prodigious, that is to say, which is completely abhorrent and against Nature. . . .
It is certain that most often these monstrous and prodigious creatures proceed from the judgment of God, who permits fathers and mothers to produce such abominations from the disorder that they make in copulation, like brutish beasts. . . . Similarly, Moses forbids such coupling in Leviticus (Chapter 16) (fig. 2.3).[30]
The creatures discussed in this chapter are produced by the natural cause of the fusing together of strange species, but, more important,
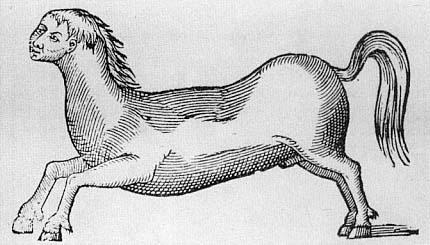
Fig. 2.3.
A colt with a man's head.
their, so to speak, first cause is God's wrath at the copulation between human beings and other species, a practice that is explicitly forbidden in Leviticus. The result is not only a monster but a prodigy, a creature that is contrary to nature and that is described as completely abhorrent.
If we turn to the chapter that treats the natural cause of the mixture or mingling of seed, we find Paré endorsing the principle that nature always strives to create its likeness; since nature always preserves its kind and species, when two animals of different species copulate, the result will be a creature that combines the form of both of the species.[31] The kind of naturalistic explanation exhibited in this chapter is, however, framed by crucial opening and closing paragraphs, which I quote at length. The chapter begins with this statement:
There are monsters that are born with a form that is half-animal and half-human . . . which are produced by sodomists and atheists who join together, and break out of their bounds contrary to nature, with animals, and from this are born several monsters that are hideous and very scandalous to look at or speak about. Yet the disgrace lies in the deed and not in words; and it is, when it is done, a very unfortunate and abominable thing, and a great horror for a man or woman to mix with and copulate with brute animals; and as a result some are born half-men and half-animals (figs. 2.4, 2.5).[32]
The chapter closes with this:
Now I shal refrain from writing here about several other monsters engendered from such grist, together with their portraits, which are so hideous and abominable, not only to see, but also to hear tell of, that, due to their
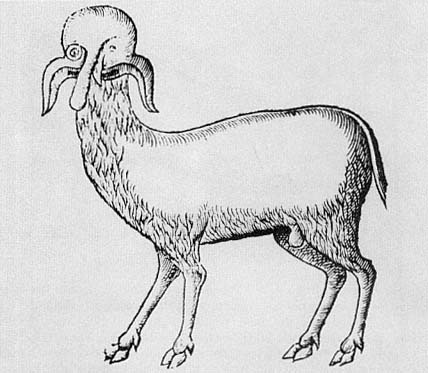
Fig. 2.4.
A monstrous lamb.
great loathsomeness I have neither wanted to relate them nor have them portrayed. For (as Boaistuau says, after having related several sacred and profane stories, which are all filled with grievous punishments for lechers) what can atheists and sodomists expect, who (as I said above) couple against God and Nature with brute animals?[33]
What I want to isolate is the conjunction of God's wrath at human disobedience of his laws (a supernatural cause) with the production of a creature contrary to nature, a prodigy, the reaction to which is horror; and, finally, I want to emphasized that the prime example for Paré of such human disobedience is bestiality. These features are in effect Paré's analogue to Aquinas's discussion in the Summa Theologica . For Thomas, there is a distinct category of lust, worse in kind than other species of lust, namely, just contrary to nature (remember that prodigies, being completely against nature, are worse in kind than monsters, being only outside the course of nature), the most grievous example of which is bestiality; moreover, when such sins are committed, an injury is done to God. Paré physicalizes this framework of concepts by exhibiting the consequence of such an injury to God; the resulting bestial creature is a sym-

Fig. 2.5.
A child, half dog.
bolic representation of God's wrath, and the reaction of horror we have to such hideous creatures is intended to remind us of, and to impress upon us, the horror of the sin itself. Thus, the special viciousness of sins contrary to nature extends to the creatures produced by these sins. Paré reserves his most charged language—horror, horrible, hideous, loathsome, abominable—for these creatures and the sins they represent.
The link between moral disorder and the disorder of nature was a constant theme during this period. It was widely believed that evil committed on earth could leave its mark on the structure of the human body.[34] And the way in which the physical form of the body gave rise to moral and theological questions went far beyond the case of prodigies. The issue of monstrous births as a whole raised practical problems for priests, since they had to decide whether any particular monstrous child was human, and so whether it should be baptized or not. There were, of course, disagreements about how to make these determinations, but the form of the body served as a guide to theological resolution. The kind of reasoning employed is well represented by Guido of Mont Rocher's Manipulus Curatorum Officia Sacerdotus of 1480:
But what if there is a single monster which has two bodies joined together: ought it to be baptized as one person or as two? I say that since baptism is made according to the soul and not according to the body, howsoever there be two bodies, if these is only one soul, then it ought to be baptized as one person. But if these are two souls, it ought to be baptized as two persons. But how is it to be known if there be one or two? I say that if there be two bodies, there are two souls. But if there is one body, there is one soul. And for this reason it may be supposed that if there be two chests and two heads there are two souls. If, however, there be one chest and one head, however much the other members be doubled, there is only one soul.[35]
I mention this example to indicate that Paré's use of the body as a moral and theological cipher is only a special instance, and not an entirely distinctive one, of a much more general mentalité.
What is most remarkable aobut Paré's book is that when he confines himself to purely natural causes, he employs the concept of monster exclusively (phenomena outside the course of nature) and not the concept of prodigy. Furthermore, the experience of horrror is absent from his descriptions. Horror is appropriate only if occasioned by a normative cause, the violation of some norm, as when the human will acts contrary to the divine will. The chapter that immediately follows Paré's discussion of the wrath of God concerns monsters caused by too great a quantity of seed. Compare its opening language with the language of the previous chapter already quoted.
On the generation of monsters, Hippocrates says that if there is too great an abundance of matter, multiple births will occur, or else a monstrous child having superfluous and useless parts, such as two heads, four arms, four legs, six digits on the hands and feet, or other things. And on the contrary, if the seed is lacking in quantity, some member will be lacking, [such] as feet or head, or [having] some other part missing (figs. 2.6, 2.7).[36]
Even Paré's discussion of hermaphrodites in chapter 6 bears no trace of horror, and we see that their formation is due entirely to natural causes,
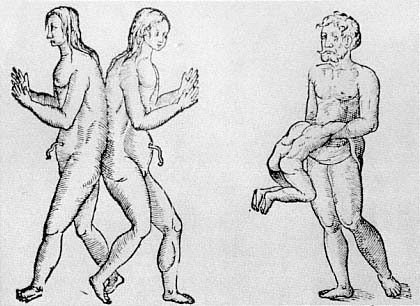
Fig. 2.6.
Examples of too great a quantity of seed.
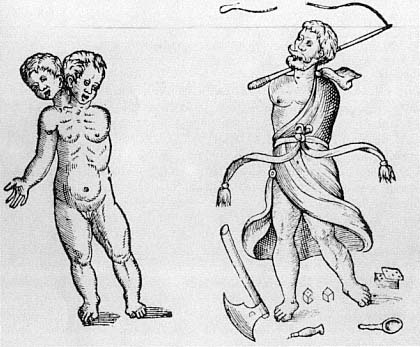
Fig. 2.7.
Examples of lack in the quantity of seed.
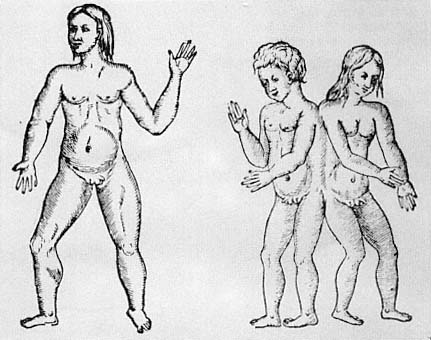
Fig. 2.8.
Hermaphrodites.
with no admixture of willful violation of a norm (fig. 2.8). Hermaphrodites are monsters, not prodigies, naturally explicable and normatively neutral.
If we read Paré's treatise chapter by chapter, we find that horror is a normative reaction, a reaction engendered by a violation of a specific kind of norm. When causal knowledge, that is, knowledge of the natural causes, is produced to explain a monster, the effect of such explanation is to displace horror, to alter our experiences of the phenomenon with which we are confronted. Horror is linked to Paré's discussion of supernatural causes because the issue in these discussions is always the normative relation between the divine and human wills. A horrible prodigy is produced when the human will acts contrary to nature, contrary to the divine will, and so when this contrariness (as Aquinas makes conceptually articulate and as is reflected in Paré) involves the thwarting of a very particular kind of norm. I see no reason to doubt the accuracy of Paré's descriptions, of where and when he experienced horror, especially because this kind of description is confirmed in so many other treatises.[37] It strikes me as no odder that Paré and his contemporaries would experi-
ence horror only when confronted by a prodigy, by some especially vicious normative violation, than that the Israelites of the Old Testament would come to experience horror at the seemingly heterogeneous group of phenomena called "abominations." And the inverse relationship between horror and causal explanation is the other side of the similar relationship between wonder and causal explanation. A sense of wonder was the appropriate reaction to the production of a miracle, just as horror was the appropriate reaction to the production of a prodigy. Lorraine Daston has argued, in examining the decline of miracles and the sensibility of wonder, that "it was axiomatic in the psychology of miracles that causal knowledge drove out wonder, and in the seventeenth century the converse was also emphasized: wonder drove out causal knowledge."[38] The psychology of miracles and the psychology of prodigies were phenomenologically and analytically akin to each other.
In his chapter on the mixing and mingling of seed and the hideous monsters that result from bestiality (fig. 2.9), Paré describes a man-pig, a creature born in Brussels in 1564, having a man's face, arms, and hands, and so representing humanity above the shoulders, and having the hind legs and hindquarters of a swine and the genitals of a sow (fig. 2.10). This man-pig was one of a little of six pigs and, according to Paré, "it nursed like the others and lived two days: then it was killed along with the sow on account of the horror the people had of it."[39] As one would expect from what I have argued, horror was in fact the reaction triggered by this man-pig, and it was so consuming as to push the people to kill both the sow and her monstrous offspring.
In 1699, Edward Tyson, a fellow of the Royal Society and Royal College of Physicians, communicated a report, published in the Philosophical Transactions of the Royal Society, entitled "A Relation of two Monstrous Pigs, with the Resemblance of Human Faces, and two young Turkeys joined by the Breast." Tyson announces his intention at the start:
By the description of the following monsters I design to prove that the distortion of the parts of a fetus may occasion it to represent the figure of different animals, without any real coition betwixt the two species.[40]
He proceeds to describe, in much detail, a so-called man-pig, discovered at Staffordshire in 1699. His article contains no evidence of horror, disgust, dread, or any related emotion. As he continues, it becomes clear that his description of the seemingly human face of the pig is meant to show that it is the result of some depression of the pig's face, caused by a compression of the womb or by the pressure of the other pigs in the same part of the womb. No reference to bestiality is necessary to understand the production of this creature, and no horror is, or should be occasioned by it. Tyson mentions the case of a man-pig reported by Paré,
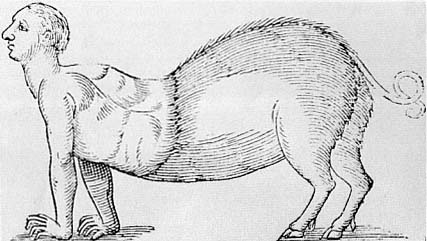
Fig. 2.9.
A monster, half man and half swine.
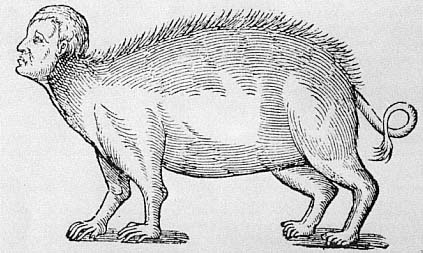
Fig. 2.10.
A pig, having the head, feet, and hands of a man, and the rest of a pig.
the very case I have quoted, and is content to point out some differences between Paré's case and his, for example, that his man-pig did not possess human hands. Tyson is cautious about whether recourse to bestiality is ever required to explain such monsters, but the main thrust of his article is to show that causal explanations of the kind he has produced have a much greater explanatory relevance than has often been recognized. His attitude stands at a great distance from Paré's, and it is exem-
plified by his remark, made when discussing other reported cases of monstrous pigs, "I believe either fiction, or want of observation has made more monsters than nature ever produced"[41] —sometimes almost employing the concept of monster as if monsters were thought to be creatures contrary to nature, whereas the whole point of his communication has been to show that they result from abnormal deformations due to natural causes.
The displacement of horror as a result of causal explanation, as though knowing the cause of a monster calms the horror we might feel, can also be seen in the case of John Merrick, the so-called Elephant Man, and such displacement operates in one and the same individual, namely, Merrick's physician, Frederick Traves (figs. 2.11, 2.12). In the medical reports submitted to the Pathological Society of London, words such as "deformity," "abnormality," "remarkable," "extraordinary," and "grossly" describe Merrick's condition. The reports do not convey an experience of horror but rather an impression of how extreme Merrick's deformities are, and, because of that extremity, they indicate the immense medical interest of his condition. However, when we read Treves's memoir and he describes his and others' first encounters with the Elephant Man, the mood is completely different. Here we find words and phrases such as "repellent," "fright," "aversion," "a frightful creature that could only have been possible in a nightmare," "the most disgusting specimen of humanity that I have ever seen," "the loathing insinuation of a man being changed into an animal," and "everyone he met confronted him with a look of horror and disgust."[42] (See fig. 2.13.) It is as though we can describe Treves's emotional history by saying that when he attends to the complicated causal etiology of Merrick's condition, he can transform his own reaction from one of horror and disgust to pity and, eventually, sympathy. We often assume that the appellation "Elephant Man" derives from the fact that Merrick was covered with papillomatous growths, a derivation of this name reported in one of the medical communications. And certainly this appearance could have accounted for that title. But it is easy to forget that this is not the official reason that Merrick himself gave for his being called the Elephant Man. He reported that shortly before his birth, his mother was knocked down by a circus elephant and that this accident, with its horrifying consequences, was the source of the label, "Elephant Man." It is perfectly evident that this story conceals, and not very well, the fantasy of bestiality, and it is exactly this fantasy that is embedded in Traves's memoir when he speaks of "the loathing insinuation of a man being changed into an animal."
Although the adjective "abominable" occurs frequently in discussions of monsters and prodigies, I will not insist here on the obvious differences between this use of the term and the concept of abomination in the
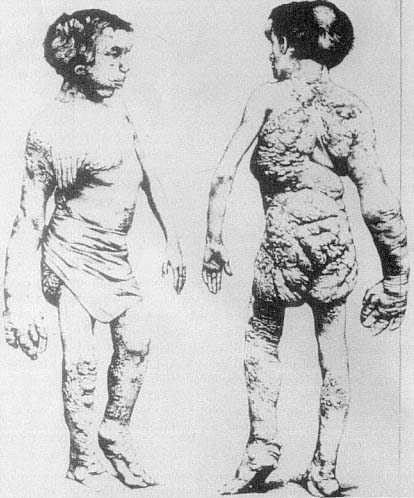
Fig. 2.11.
John Merrick, 1884–85.
Old Testament. The use of "abominable" to describe prodigies remains inextricably linked to horror, as I have argued; but the doctrine of natural law, absent from the Old Testament, decisively alters one feature of the biblical conception. A study of the relevant biblical passages would show that it is primarily one uniquely specified people who, because of their special relation to God, feel horror at abominations. But in the texts I have discussed, it is rather as though the horror of sins contrary to nature, and of the products that result from them, is experienced by all human beings qua rational beings. For the use of natural reason alone is sufficient to grasp the viciousness of sins contrary to nature, and bestial-
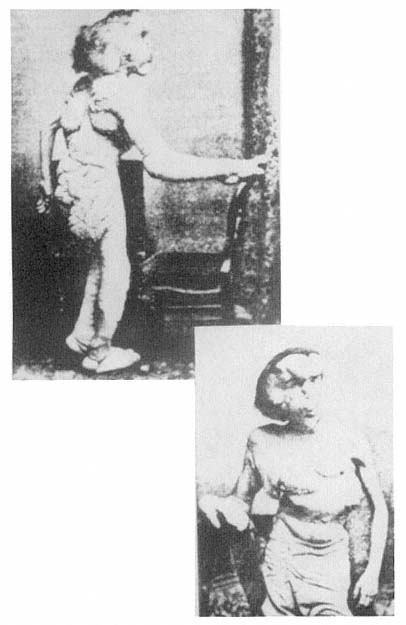
Fig. 2.12.
John Merrick.
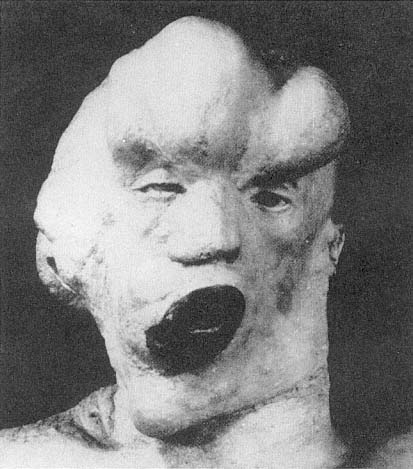
Fig. 2.13.
Postmortem cast of the head and neck of John Merrick.
ity, for example, is a violation of natural law, which requires no special act of divine revelation to be known but is nothing else than the rational creature's participation in God's eternal law.[43] So every human being ought to experience horror at that which he knows, as a rational being, to be contrary to nature. In this context, the doctrine of natural law helped to conceal the recognition that horror is a cultural and historical product and not demanded by reason alone, a fact that is more easily recognized in the pertinent biblical texts. Since horror came to be enmeshed in the framework of natural law and natural reason, prodigies, and the wrath of God, could be described in a way that was intended to represent the
experience of every human being, not simply the experience of a culturally specific group. Objects of horror could now directly appear to be naturally horrifying.
As I have already shown, bestiality, the worst of the sins contrary to nature, exhibited its viciousness in the very structure of the human body itself, in the creatures produced by the willful violation of God's natural law. But this configuration, whereby a certain kind of undermining of norms was exhibited in the effects of physical pathology, was not restricted only to this one form of lust contrary to nature. Eighteenth- and early nineteenth-century treatises on onanism reproduce this same pattern of concepts; self-abuse, another one of Aquinas's sins contrary to nature, ravages the physical structure of the body, producing, among its effects, acute stomach pains; habitual vomiting that resists all remedies during the time this vicious habit is continued; a dry cough; a hoarse, weak voice; a great loss of strength; paleness; sometimes a light but continuous yellowness; pimples, particularly on the forehead, temples, and near the nose; considerable emaciation; an astonishing sensibility to changes in the weather; an enfeeblement of sight sometimes leading to blindness; a considerable diminution of all mental faculties often culminating in insanity; and even death (fig. 2.14).[44] Indeed, this relationship between the viciousness of the sin and the pathology of the body even gave rise to a genre of autopsy report, in which the autopsy of a masturbator would reveal that the effects of this loathsome habit had penetrated within the body itself, affecting the internal organs no less than the external appearance.[45] In Tissot's L'Onanisme, Dissertation sur les maladies produites par la masturbation, we find the same kind of terminology and sensibility that accompanies Renaissance descriptions of prodigies. Tissot opens his discussion of cases, with which he has had firsthand experience, with the following preamble:
My first case presents a scene which is dreadful. I was myself frightened the first time I saw the unfortunate patient who is its subject. I then felt, more than I ever had before, the necessity of showing young people all the horrors of the abyss into which they voluntarily plunge themselves.[46]
And he invokes the idea of masturbation as contrary to nature in strategically central passages.[47]
It is often said that Tissot's treatise is the first scientific study of masturbation, and his book is engulfed by medical terminology and punctuated by attempts to give physiological explanations of the pathological effects provoked by masturbation. But it is just as evident that his book remains firmly placed within a tradition of moral theology, which begins with a conception of masturbation as an especially vicious kind of lust. It produces mental and physical disease and disorder, but even in the scien-

Fig. 2.14.
Death by masturbation.
tific treatments inaugurated by Tissot, it remains a vicious habit, not itself a disease but a moral crime against God and nature. Tissot begins his book with the claim, which he says that physicians of all ages unanimously believe, that the loss of one ounce of seminal fluid enfeebles one more than the loss of forty ounces of blood.[48] He immediately recognizes that he must then explain why the loss of a great quantity of seminal fluid by masturbation, by means contrary to nature, produces diseases so much more terrible than the loss of an equal quantity of seminal fluid by natural intercourse. When he offers an explanation, in Article II, Section 8 of his book, he attempts to frame it in terms of purely physical causes, the mechanical laws of the body and of its union with the mind. But try as he might, he cannot help but conclude this section by reintroducing the claim that masturbators "find themselves guilty of a crime for which divine justice cannot suspend punishment."[49]
Theorists of sodomy also exploited this same kind of connection between normative taint and physical deformation. The normative origin of attitudes toward sodomy is contained not only in the very word, with its reference to the episode of Sodom and Gomorrah in Genesis, but also in the emergence of other words to refer to the same practices. For instance, buggery derives from the French bougrerie, a word that refers to a Manichaean sect that arose at Constantinople in the ninth century, and
which recognized a sort of pontiff who resided in Bulgaria. Thus, to be a bougre meant that one was a participant in heresy, and there is no reason to believe that this heretical sect had any special proclivity toward sodomy. However, eventually, the accusation of bougrerie came to be identified with an accusation of sodomy, and the link to heresy became submerged.[50] Moreover, in French, the phrase "change of religion" could be used to describe pederasty; to become a pederast was to change religion (changer de religion ).[51] Both sex and religion have their orthodoxies, their heresies, their apostasies—their nomrative paths and deviations.
Even when the theological underpinnings of the concept of sodomy receded into the background, its normative content and origin was always close at hand. Ambroise Tardieu, whose enormously influential Étude médico-légale sur les attentats aux moeurs was first published in 1857, devotes about one-third of this book to a discussion of pederasty and sodomy. Tardieu restricts the term "pederasty" to the love of young boys, whereas the more general term "sodomy" is reserved for "acts contrary to nature, considered in themselves, and without reference to the sex of the individuals between whom the culpable relations are established."[52] Most of the cases of sodomy Tardieu describes concern either male/male anal intercourse or male/female anal intercourse. The fact that he repeatedly characterizes these acts as contrary to nature indicates the normative tradition into which his work fits. Although Tardieu acknowledges that madness may accompany pederasty and sodomy, he wishes to make certain that these acts escape "neither the responsibility of conscience, the just severity of law, nor, above all, the contempt of decent people."[53] He is aware that the "shame and disgust"[54] that these acts inspire have often constrained the reports of observers, and his book is intended to remedy this lack, and in extraordinary detail.
Much of Tardieu's discussion of pederasty and sodomy is concerned with the physical signs that permit one to recognize that these activities have transpired, with the material traces left by these vices in the structure of the organs. Tardieu believed that an exhaustive discussion of these signs is necessary if legal medicine was going to be able to determine with assurance whether such acts contrary to nature and public morality had taken place. He describes the deformations of the anus that result from the habit of passive sodomy, a topic that had already received much discussion in the French and German medicolegal literature. But he goes on to describe the signs of active pederasty, signs left on the viril member itself, which he claims have been completely ignored in previous treatises. Changes in the dimension and form of the penis are the most reliable indications of active sodomy and pederasty. The active sodomist has a penis that is either very thin or very voluminous. The excessively voluminous penis is analogized to "the snout of certain animals,"[55] while
Tardieu describes the much more common, excessively thin penis of active sodomists in the following remarkable way:
In the case where it is small and thin, it grows considerably thinner from the base to the tip, which is very tapered, like the finger of a glove, and recalls completely the canum more .[56]
To confirm his general observations, he reports the physical conformation of the penises of many active sodomists:
Having made him completely undress, we can verify that the viril member, very long and voluminous, presents at its tip a characteristic elongation and tapering that gives to the gland the almost pointed form of the penis of a dog.[57]
Another of Tardieu's active sodomists has a penis that "simulates exactly the form of the penis of a pure-bred dog."[58] As if to confirm that sodomy is contrary to nature and God, the relevant parts of the human body are transformed by this activity so that they come to resemble the bodily parts of a dog. What could be more horrifying than the moral and physical transformation of the human into a beast, a man-dog produced no longer by bestiality but by the disgusting practice of sodomy. Long after the classical discussions of prodigies, the category of the contrary to nature continued to mark out one fundamental domain of horror.
By the late nineteenth century, the experiences provoked by so-called freak shows already contrasted with the horror of the contrary to nature. Rather than exhibiting the physical consequences of normative deviation, the freaks exhibited in sideshows and circuses were intended to amuse, entertain, and divert their audiences. In general,
the urban workers who came to stare at freaks were by and large an unsophisticated audience in search of cheap and simple entertainment. . . . In the early 1870s William Cameron Coup had introduced a two-ring concept while working with Barnum and by 1885 most shows revolved around a multiple ring system. The result was a drift toward glamour and spectacle as the basic product of the big shows. The tendency was well developed by the early nineties and brought specific changes to the exhibits. Contrasts of scale—fat ladies and living skeletons, giants and dwarfs—and exhibits involving internal contrasts—bearded ladies, hermaphroditic men and ladies toying with snakes—began to displace the more repulsive exhibits. As the shows were freighted with fewer mutilated horrors they became less emotionally loaded and less complex as experiences.[59]
It should be noted that part of the purpose of the multiple-ring circus would have been defeated by the displaying of horrors. For if having more than one ring was intended to get the spectators to look from exhibit to exhibit, to gaze periodically and repeatedly at each of the

Fig. 2.15.
Fred Wilson, the Lobster Boy.
rings, to experience the circuis in all of its diversity, then the exhibition of a horrifying object would have tended to thwart this experience. The experience of horror disposes us to fix on its object, unable to avert our gaze, fascinated as well as repulsed, blocking out virtually everything but the object before our eyes. Thus, horror is incompatible with the glamour, spectactle, and variety that is inherent in the multiple-ring circus.

Fig. 2.16.
Avery Childs, the Frog Boy.
The modern circuis has to be set up so that no single exhibit so predominated that the many rings were, in effect, reduced to one.
Even if we put aside the fact that the categories of freaks and prodigies were by no means composed of the same specimens, we can see how different this experience of freaks was by examining photograph of them. Charles Eisenmann was a Bowery photographer who took many portraits of freaks during the late nineteenth century. Some of these photographs represent characters that are half-human and half-animal and so, at least in this respect, can be thought of as successors to the medieval and Renaissance prodigies produced by bestiality. But these photographs exhibit no indication of horror. Avery Childs, the Frog Boy, is evocative and amusingly photographed but no more horrifying than a contortionist, his slippers emphasizing that he is more human than frog (fig. 2.15). Indeed, these photographs insist on the humanity of their subjects, precisely the opposite of Paré's discussions, which highlight the

Fig. 2.17.
Jo Jo, the Russian Dog Face Boy.
bestiality of prodigies. Fred Wilson, the Lobster Boy, suffers from a serious congenital deformity, but dressed in his Sunday best, with his hair neatly combed, one is drawn as much to his human face as to his supposed lobster claws (fig. 2.16). And even Jo Jo, the Russian Dog Face Boy, one of Barnum's most famous attractions, wears a fringed valour suit and sports that great symbol of Western civilization, the watch chain (fig. 2.17). Furthermore, his right hand bears a ring, and his left hand is neatly placed on his knee. And he poses with a gun, as if to suggest that he is not an animal to be hunted but can himself participate in the all too human activity of the hunt. Horror at the prodigious, amusement by the freak—the history of monsters encodes a complicated and changing history of emotion, one that helps to reveal to us the structures and limits of the human community.
Three—
The Animal Connection*
Harriet Ritvo
The dichotomy between humans and animals—or man and beast, as it used to be called—is so old and automatic that we scarcely notice it. It was enshrined near the beginning of our tradition, in the second chapter of Genesis, when God presented the animals to Adam one by one, in the vain hope that one of them would prove a fit helpmeet for him. In the end, of course, none of them would do, and God had to provide Adam with a creature more like himself.[1] At least since then, the notion that animals are radically other, on the far side of an unbridgeable chasm constructed by their lack of either reason or soul, has been a constant feature of Western theology and philosophy.[2] It has completely overshadowed the most readily available alternative, which would define human beings as one animal kind among many others.
And the advent of modern science has not made much difference. Most scientific research about animals has been founded on the assumption that they constitute a distinct class to which human beings do not belong. Thus, eighteenth- and nineteenth-century zoologists were confident that "the prerogative of reason," which animals lacked, distinguished humanity "as . . . intended for higher duties, and a more exalted destiny."[3] This confidence insulated them from the implications of such disquieting recognitions as the following, taken from an early Victorian zoological popularizer: "When we turn our attention to Mammalia . . . we find some startling us by forms and actios so much resembling our own, as to excite unpleasant comparisons."[4] Nor did Charled Darwin's formulations necessarily change things. The gap between humans and animals remained axiomatic even, or perhaps especially, in fields like comparative psychology, which focused on the kinds of intellectual and emotional qualities that were also assumed to distinguish us as a species.
George Romanes, a pioneer in such research and the friend and protégé of Darwin, tried to quantify this discontinuity in a book entitled Mental Evolution in Animals, published in 1883. It included a graphic scale of emotional and intellectual development, presented as a ladder with fifty steps. Civilized human adults, capable of "reflection and selfconscious thought," were at the top. Then there was a large hiatus. The highest animals were anthropoid apes and dogs, which Romanes considered capable of "indefinite morality" as well as shame, remorse, deceitfulness, and a sense of the ludicrous. They occupied step 28, along with human infants of fifteen months. Close behind them, on step 25, were birds, which could recognize pictures, understand word, and feel terror; and they were followed on step 24 by bees and ants, which could communicate ideas and feel sympathy.[5] The details of this schematization sound quaint now, but its underlying taxonomy, which defines humans and animals as separate and equivalent categories on the basis of their intellectual, spiritual, and emotional capacities, continued to determine the course of research on animal behavior for the succeeding century. As Donald Griffin, a contemporary critic of this taxonomy, has pointed out. "Throughout our educational system students are taught that it is unscientific to ask what an animal thinks or feels . . . [and] field naturalists are reluctant to report or analyze observations of animal behavior that suggest conscious awareness . . . lest they be judged uncritical, or even ostracized from the scientific community."[6]
Although the dichotomy between humans and animals is an intellectual construction that begs a very important question, we are not apt to see it in that light unless it is challenged. And serious challenges have proved difficult to mount. Those who have based their thinking on the uniqueness of our species (i.e., its uniqueness in a different sense than that in which every species is unique) have often resisted even the attempt to make the dichotomy controversial. The scientific consensus cited by Griffin exemplifies entrenched institutional reluctance to acknowledge that an alternative taxonomy might be possible. An analogous refusal by philosopher Robert Nozick structured his review, which appeared several years ago in the New York Times Book Review, of Tom Regan's The Case for Animal Rights . Instead of grappling seriously with Regan's carefully worked out and elaborately researched argument, Nozick simply dismissed it by asserting that animals are not human and therefore cannot possibly have any rights. That is, he claimed that Regan had made a crippling category mistake by failing to recognize the insuperable barrier that separated humans from all other creatures and that it was therefore not necessary to think seriously about anything else that he said.[7] Such views are not confined to scholars and scientists; so, despite its evasiveness, Nozick's stratagem is unlikely to have bothered
many of his readers. Recent research suggests that most ordinary Americans explicitly endorse the dichotomy that Nozick postulates, whatever else they may think or feel about animals, for example, whether or not they like them, or whether they wish to protect them or to exploit them.[8]
But this repeatedly avowd taxonomy is not the whole story, either about the relationship of human beings to other species or about the way that people have perceived and interpreted that relationship. There are other indexes of belief and understanding than explicit declarations. In the case of other animals, and especially the mammalian species that human beings resemble most closely, the explicit denial of continuity may paradoxically have freed people to articulate, in a veiled and unselfconscious way, their competing sense of similarity and connection. A lot of evidence suggests that when people are not trying to deny that humans and animals belong to the same moral and intellectual continuum, they automatically assume that they do. Discourses that seem to refer exclusively to animals are frequently shaped by cultural constructions clearly derived from human society, even in the scientific and technological fields where it might seem that such constructions would be counterproductive, out of place, and easy to identify and discard. The consequences of this unacknowledged connection have often been enormous, even in the behavioral sciences most strongly committed to reinforcing the dichotomy between humans and animals. Thus, it is no accident that the baboon studies published by S. L. Washburn and Irven DeVore in the 1950s and 1960s stressed the importance of male dominance hierarchies. Analogously, the research undertaken by the increasing number of female primatologists in the past two decades has emphasized the extent to which female primates aggressively manage their own reproductive careers, radically revising earlier characterizations of them as sexually passive and even "coy."[9]
thus animal-related discourse has often functioned as an extended, if unacknowledged metonymy, offering participants a concealed forum for the expression of opinions and worries imported from the human cultural arena. Indeed, much of what people—experts of one sort or another—have said about animals can only be explained in this context. Thus, the foregoing examples from the recent history of primatology suggest how social or political ideology can determine the research agenda of scientists. But these examples may seem too easy. After all, despite the explicit professional commitment of primatologists not to anthropomorphize the animals they study, those creatures are of special interest exactly because of their closeness to humankind. They are obvious targets for projection, as are the extinct pongide and hominids whose fossil remains are interpreted by students of human origins.[10]
It is, however, possible to find evidence that the same intellectual and
cultural preconceptions shape discourses that, on the face of it, look much less promising. One such discourse informs the literature of animal breeding that emerged in late eighteenth-century Britain and developed and flourished throughout the next century. The ostensible subjects of this discourse—horses, cattle, sheep, pigs, dogs, and cats—share far fewer characteristics with human beings than do apes, monkeys, and australopithecines; its participants were concerned with practical results, rather than with anything so abstract and potentially tendentious as the increase of knowledge or the development of theory.
The flavor of this literature—both its tone and its content—is more easily demonstrated by excerpts than by general characterization. The following selections touch in various ways on the relations between the sexes, a subject that was crucial to successful animal husbandry; they also make assertions that are difficult to understand with reference only to the exigencies of stock breeding. In 1828, the author of an article that appeared in the Farrier and Naturalist, a livestock-oriented journal, asked, "What is . . . the part of the female in the great act of reproduction?" He answered his own question, "When the male predominates by his vigour, his constitution, and his health, she is limited, in some measure, to perform the same office that the earth does for vegetables . . . nothing more than a receptacle, in which are deposited the seeds of generation."[11] A few years later, William Youatt, the most distinguished British veterinarian of the early Victorian period and a prolific writer on domestic animals, recounted the following story as an illustration of the need to control the imagination of "even so dull a beast as the cow": a certain cow "chanced to come in season, while pasturing on a field . . . out of which an ox jumped, and went with the cow, until she was brought home to the bull. The ox was white, with black spots, and horned. Mr. Mustard [the owner of the cow] had not a horned beast in his possession, nor one with any white on it. Nevertheless, the produce of the following spring was a black and white calf with horns."[12] Early in the twentieth century, Judith Neville Lytton, a prominent if iconoclastic and combative member of the toy dog fancy, suggested the following remedy for barreness in prize bitches: "In desperate cases . . . try the old . . . recipe of breeding to a thorough cur. . . . If the bitch breeds to this connection . . . the next time the bitch is put to a thoroughbred dog she will almost certainly breed to him. . . . The more . . . highly bred the bitch is, the more likely this is to succeed."[13]
Each of these statements depended on assumptions not only obviously false in the light of modern science but also subject to persuasive contradiction on the basis of accumulated practical experience that was widely available in the nineteenth century. Each statement uses, applied to the activities of domestic animals, language ordinarily reserved to describe
human social intercourse. Why, then, did experts—the very people who should have access to the most enlightened thought and the widest information—hold these opinions (or believe these facts, as they would have put it), and why did they express them in this way? It is likely that both the form and the content of these excerpts were determined by the exigencies of human gender stereotypes rather than by those of applied biology.
This was the case even though animal husbandry was a quintessentially earthbound pursuit, constrained by physicality and detail on every side. It had no obvious connection to the stakes at issue in any arena of human social discourse, including that of gender. Certainly, the techniques used to breed animals were very different from those used to breed people. The results of breeding were highly concrete, and they were usually presented to the general public in the stripped-down terms of cash value. Given all this, it would be reasonable to assume that breeders' understanding of their craft would be structured by empirical rather than rhetorical considerations. Yet what they said about their cattle, sheep, and dogs was strongly conditioned by their views about the nature of human beings, especially women.
This assertion may seem particularly surprising because, by the nineteenth century, the breeding of pedigreed animals had become a highly technical and specialized endeavor, whether it was carried on by professional agriculturalists interested in producing improved farm livestock or by self-professed amateurs who concentrated on dogs, cats, and such smaller animals as guinea pigs, rabbits, and poultry. It was crudely materialistic in its explicit aims. Since the primary goal of all breeders was to produce superior young animals, the crucial focus of their attention was the selection of healthy and appropriately endowed parents for the new generation. Several factors encouraged them to be as pragmatic as possible in their matchmaking decisions.
In the first place, mistakes were easy to spot. Stringent standards existed for almost every king of animal that was frequently bred, and these standards were widely disseminated in handbooks, prints, and periodicals and vigorously enforced by show judges and by the marketplace in which animals were brought and sold by knolwedgeable fanciers. (There were occasional exceptions to this rule of consensus and conformity, the most notable being the pig, which seemed so gross and amorphous that breeders had trouble figuring out what an ideal animal should be like.)[14] These frequently reiterated standards meant that the inferiority of the offspring of ill-considered pairings would sooner or later become obvious—perhaps at birth, but certainly by the time they reached maturity. In addition, breeding was an expensive pursuit—absolutely expensive in the case of the larger animals, which could cost
hundreds and even thousands of pounds to purchase and then to maintain in an appropriate style, and relatively expensive in the case of the smaller ones. Any pregnancy risked the life of the mother, and each successful pregnancy consumed a significant portion of her reproductive potential. Owners of valuable female animals had to expend this limited resource very carefully.
In the course of the eighteenth and nineteenth centuries, the expertise of breeders produced some impressive results. (This seems true even though recent historians have persuasively modified the most extravagant claims made by participants in the Agricultural Revolution and their subsequent admirers.)[15] The earlier and most celebrated achievement of English animal breeding was the modern thoroughbred racehorse, which appeared toward the beginning of the eighteenth century as the result of an infusion of Arabian blood into native English equine stock. The merit of such horses was easily measured on the track. By the middle of the eighteenth century, agriculturalists were applying the techniques developed by racehorse breeders to farm livestock, with consequent impressive increases in the size, most notably, of cattle and sheep but also of pigs and draft horses. And the nineteenth century saw a related explosion in the diversity of fancy animals. Most modern dog breeds originated then, despite the calims of greater qntiquity made by some aficionados; the same was true for cats, rodents, and the diverse starins of pigeons that Darwin studied.
This record of serious animal husbandry in eighteenth- and nineteenth-century Britain thus seems to be one of straightforward, quantifiable, pragmatically oriented achievement. New methods were developed—albeit mostly through trail and error, rather than as a result of systematic theorizing—and carefully applied, with predictably impressive results. This was the way the master breeders themselves understood their accomplishments, as documented in their published reflections on their craft. Such reflections usually took one of two forms: advice to beginners or records of particular successes for the edification of posterity. Authors working in both genres suggested that the complex procedures they described could be rather mechanically applied either to the improvement of whole breeds by those at the forefront of husbandry or to the imitation of such results by breeders content to follow modestly in paths blazed by others. Thus, one early Victorian manual for sheep breeders confidently associated the method with the result, asserting that "there cannot be a more certain sign of the rapid advanced of a people in civilization and prosperity, than increasing attention to the improvement of livestock"; in a related vein, an earlier agricultural treatise had assured readers that "the perfecting stock already well-bred is a pleasant, short and easy task."[16] At the end
of the century, the author of a handbook for cat fanciers similarly suggested that good results would follow the methodical application of expertise: "[mating] requires . . . careful consideration, and . . . experience and theory join hands, while the knowledge of the naturalist and fancier is of . . . superlative value."[17] Pedigree charts, the ubiquitous schematic representations of the results of animal breeding, also corroborated this rather mechanical sense of what the enterprise involved.
When their discussions of animal breeding became more specific, however, the experts tended to retreat from their assertive self-confidence. Neither method or knowledge, even when operating on cats of impeccable pedigree and robust health, could assure "anything like certainty," according to the expert just cited.[18] Manuals for breeders of cattle, sheep, and horses often warned novices not to attempt to produce the kinds of animals that won prizes at national shows, because of the difficulty, risk, and expense involved. Thus, when closely scrutinized, animal breeding no longer seemed merely a mechanical, if complex, technical procedure but was implicitly redefined as a more ambiguous and impressionistic activity. And the more precisely the instructions were articulated, the more confusing they became. Often experts raised issues or offered advice that was irrelevant to the achievement of their stated aims, or even counterproductive. Old and widely recognized canards were ritually chewed over for decades after they had been persuasively discredited.
An explanation might strees the unimaginative and derivative nature of many of these works, which occupied the borderline between technical and popular writing, or it might focus on the conservatism inherent in many fields of applied and popularized technology. And there is doubtless some truth to both of these possibilities. But the curious or anomalous elements in the discourse of animal breeding can be more fully explained if that discourse is also viewed as an arena for the discussion of human gender issues. In a way, it was an extremely obvious focus for each concerns. After all, the central task of breeders was to manage sexual relations of their animals. This task often posed challenges beyond the merely intellectual problem of deciding which ones to pair up. The fact that the participants in this discourse were unaware of its double function merely allowed them to air their views and worries more directly.
In making their decisions about which animals to pair, breeders selected parents on the basis of both their individual quality (i.e., the extent to which they possessed the characteristics that were desired in their offspring) and a set of general notions about the way that the transmission of such characteristics took place. For most of the nineteenth century, there were few authoritative constraints on such ideas. Despite the
claims of scienticity that agriculturalists had made since the beginning of the eighteenth-century vogue for improvement, few of them and even fewer breeders of small animals belonged to the scientific community; the works of popular natural history that they were most likely to encounter did not deal with the questions of reproductive physiology that engaged some elite biologists. And even if breeders had been aware of the most advanced contemporary research on reproduction, they could not easily have applied its results to their enterprise.
Although it was clear to scientists, as it was to breeders, that sexual intercourse was necessary if the higher animals were to reproduce, there was no expert consensus until late in the nineteenth century about why this was so.[19] That is, the modern understanding of the balanced contribution of sperm and egg to the development of a new organism was unavailable to eighteenth- and nineteenth-century animal breeders. Without this knowledge, they were free to predict and interpret the results of their breeding ventures with reference only to their own experience. That experience was vast and, indeed, considered extremely valuable by ope-minded scientists like Charles Darwin.[20] It also turned out to include breeders' attitudes toward other people, as well as their observations of generations of animals.[21]
Many eighteenth- anc nineteenth-century theories of reproduction presented it as the gradual enlargment and development of a tiny but complete seed, but scientists who adhered to this viewpoint were divided about whether that seed was contributed by the male or the female. Animal breeders, however, were of one mind about this question. Many, like the author of the Farrier and Naturalist article quoted earlier, defined the female parent as a mere receptacle. One expert, faced with explaining why, in this case, it was not "an easy thing to produce at once very perfect animals, provided that males of the right form could be obtained," preferred not to posit the mother as a significant source of variation. Instead, he had recourse to the fact that "the offspring will, to a greater or lesser extent, partake of the form and structure of the grandparents [i.e., the grandfathers]." And even if such an absolute assertion of male dominance needed modification in view of the obvious tendency of young animals to resemble both their parents, breeding experts still reserved the more vigorous genetic role for the stud. The imagery of activity and passivity remained useful in the modified case; it suggested, for example that "the male gives the locomotive, and the female the vital organs."[22]
The preponderance of male influence never escaped the attention of writers on these subjects for long, even if they had been momentarily diverted by the need to comment on females. For example, a reminder that "without first class females the descendants will not shine . . . in the
show yard" was predictably accompanied by the acknowledgment that "it must not be forgotten that the male has most influence in breeding."[23] The only situations in which it was generally considered that the female might disproportionately determine the results of procreation were those which introduced a different, and also powerful, cultural construct. Social superiority—that is, the terms of animal husbandry, a more distinguished pedigree—might tip the scales in the direction of the female. As Youatt pointed out, the influence of "a highly bred cow will preponderate over that of the half-bred bull."[24] Since such exceptional circumstances could only result from extreme negligence or ignorance on the part of the breeder, however, they did not have to be incorporated into receive wisdom. In general, breeders were advised to proceed on the assumption that "not only . . . is the male parent . . . cpable of most speedily improved the breed of livestock . . ., but . . . the male is the parent, from motives of sense and sound polity, which we can alone look to for the improvement of our breed."[25]
Perhaps it was to maintain this strong differentation in reproductive roles that cattle show judges were adjured to assess bulls "as males and not as females" and cows "as females and not as males." The telltale signs were admittedly difficult for even first-class judges to detect—not the obvious things but subtle variations in such unlikely features as the conformation of head and horns. But the stakes were considered high enough to justify the trouble, especially where bulls were concerned. According to one veteran cattle breeder, "effeminacy in the male must be shunned as the most deadly poison."[26]
The principles that guided the production of livestock animals were routinely applied to pet species. The author of a late Citorian cat breeding manual assured his readers that "the outward characteristics are in great measure transmitted by the male cat."[27] Nor did the advance of biological knowledge necessarily shake the faith of animal breeders in their time-tested principles. Instead, as it became available, the jargon of science could be appropriated to the service of the conventional understanding of animal reproduction. Everett Millais, one of the most prominent dog fanciers of the late nineteenth century, translated it into the new terminology as follows: "that the male . . . does influence the epiblastic and mesoblastic structures largely, and all out of proportion to the female is undoubted."[28]
So powerful was the influence attributed to at least some males, it was even believed that they could determine the character of offspring in the conception of which they had had no part. That is, they might gain access to the reproductive organs through the eyes of receptive females, as well as in the ordinary way. As a result, breeders anxious to preserve the purity and the quality of their stock had to guard the minds as well as
the bodies of their impressionable female animals from such undesirable approaches. It went without saying that females would be both unable and disinclined to resist them. In short, females could not be trusted with the preservation of their own virtue, even on the level of imagination. Mr. Mustard's cow, referred to earlier, offered an extreme example of the feminine susceptibility posited by this view of relations between the sexes. The off-breed ox that jumped into her pasture when she was in heat was not even completely male—that is, he had been castrated and therefore rendered incapable of procreation—but, even so, he apparently left this make on the calf she subsequently conceived after intercourse with the properly pedigreed bull selected by her owner.
If females of a relatively stolid species were so susceptible to the influence of random males, it was not surprising that female dogs, which were considered both more intelligent and more excitable, had to be guarded still more closely. Breeders agreed that the animals they termed maiden bitches were particularly vulnerable to such external stimuli and advised that "due influence should be exercised in the thorough isolation of bitches . . . or more than a temporary evil and disappointment may occur."[29] But more experienced bitches were also at risk, and beginning breeders were warned that "even very close intimacy between a bitch during oestrum and a dog she fancies may influence the progeny, although the dog has not warded her."[30]
The struggles between bitches and breeders were described in terms that evoked stubborn daughters in romantic narratives who refused to accept their fathers' choice of suitors. Hugh Dalziel, who wrote about a variety of Victorian dog breeds, once owned a Dandie Dinmont terrier whose wayward emotions made her useless for breeding; she "became enamoured with a deerhound, and positively would not submit to be served by a dog of her own breed." Even bitches who were more compliant might defeat their owners' purposes. Delabere Blaine, sometimes known as "the father of canine pathology," had a pug bitch whose constant companion was a white spaniel. All her litters were sired by pedigreed pugs, and all consisted of undeniably pug puppies, but one in each batch was white, a color that was rare and not desirable in that breed.[31]
Recognizing that it was useless to fight against such a predilection, one prolific breeder of dogs and cats who confessed that he would allows his champion studs to serve almost any female whose owner could pay the fee made one of his rare refusals in the case of a bitch that had "already formed an attachment to a dog of a meaner breed."[32] Such inclinations were fairly common among bitches, who were likely to implement them by stubbornly resisting their owners' prudent attempts to cloister them. The author of manuals frequently warned novice breeders that bitches in heat would make unimaginably subtle and persistent attempts to es-
cape from whatever quarters they were confined in. But it was necessary to persevere in thwartint them, because the stakes at risk in the preservation of female purity were high. A match with an inappropriate partner, especially in the case of a virgin animal, was held to have consequences far beyond the issue of that particular mating, as if a female's first sexual partner in some sense established a permanent proprietorship over her reproductive capacities.
Although it was, as one late-nineteenth-century kennel expert admitted, "an exceedingly rare phenomenon," which was difficult to verify, many breeders continued to believe in what was usually called "the influence of the previous sire" but was sometimes dignified by the scientific-sounding term "telegony."[33] The midcentury author of a scientific treatise on agriculture summarized this doctrine as follows: "The effect of the first male is so great as to cause all the animals that female shall afterward give birth to, in a more or less degree, to resemble that by which the female was first impregnated." He chose to illustrate this doctrine with the story of a chestnut mare whose original mate was a quagga (a relative or variety of the zebra that became extinct late in the nineteenth century), and all of whose subsequent offspring were striped, even though all but the first resulted from her union with a black stallion.[34] The breeding literature teemed with similar examples of the dire consequences of letting females slip even once from the straight and narrow. The appearance of spotted puppies in a litter produced by two prize for terriers was explained by the fact that the sire of the bitch's previous litter had been a dalmatian.[35] Of kittens of "a good Persian sire and dam" who nevertheless appeared "remarkably poor specimens . . . what might be called half-breeds," a late Victorian cat fancier said that "I can only attribute this to the blue female having twice strayed from the paths of virtue previous to the attentions of the prize-winning Persian."[36]
Despite their frequent appearance in the breeding literature, instances of the influence of the previous sire seemed to occur only at widely spaced intervals in real life. The cases just recounted, for example, were thirdhand at best. Even in the earlier literature, therefore, authors sometimes gingerly noted that there might be ground on which to question this principle. The more scientists discovered about how reproduction actually worked, the harder it became for breeders to identify a mechanism that would account for such anomalies. They nevertheless clung tenaciously to this doctrine, perhaps because, weak though it might be as a predictor of the results of unsanctioned animal unions, it precisely expressed the metaphorical consequences of such lapses by human females. Even the most sophisticated experts hesitated to dismiss completely the possibility of influence or, as it might be more forth-
rightly expressed, contamination or infection by a female animal's first mate. At most, they might suggest that such occurrences were sufficiently rare that breeders did not need to worry about them. Thus, Neville Lytton, ordinarily impatient with the shibboleths of her fellow fanciers, conceded that "I am inclined to think [it] does occasionally happen," but she asserted that "it happens so seldom that no one has ever beey able to collect evidence enough to prove. In any case it would only affect isolated individuals, and probably only as to a single character, . . . [and so] . . . I do not think breeders need trouble themselves about so small a matter as the possible influence of a previous sire on a single puppy."[37]
In addition to jeopardizing the quality (or legitimacy) of future offspring, the tendency of female animals to follow their own sexual inclinations was perceived to pose less concrete but perhaps equally troublesome threats. Although an occasional authority might compassionately recommend indulging the desires of bitches that were "highly fed . . . [and] living luxuriously, as a means of using up their excess stock of material," an interest in sex for its own sake rather than as a means to procreation was considered an indication of depraved character.[38] The behavior of bitches, in particular, confirmed the worst male fears about female proclivities. Although a single copulation might have sufficed for pregnancy, bitches would wantonly continue to accept new partners as long as they were in heat; connected with this unseemly pursuit of pleasure was a culpable indifference to its providers. One early nineteenth-century sportsman complained that "no convincing proof of satiety is ever displayed . . . and she presents herself equally to all," with the result that the largest of her suitors "is generally brought into action." He noted with some satisfaction, however, that oversexed bitches might pay for their failure to prefer refinement to brute strength; many died while bringing forth too-large puppies.[39]
According to other authorities, however, this unmatronly behavior might harm the offspring rather than the mother. In the dog, as in some other animals that routinely give birth to multiple offspring, it is possible for a single litter to have more than one father. Breeding authorities referred to this phenomenon as 'superfoetation," a technical term that made it sound like an aberration or a disease. As even the possibility of this occurrence would jeopardize the pedigree of the resulting litter, aspiring breeders were strongly advised that "for at least a week after the bitch has visited the dog, the precautions for isolating her must not be relaxed, or all her owner's hopes may be marred."[40] But social stigma and unwelcome half-siblings were not the only ills that newly conceived pedigreed purppies might sustain as a result of their mothers' licentiousness. Dalziel suggested that during or after an unsanctioned and unneces-
sary second copulation, "excessive pain, terror, or other strong emotions, may affect the unborn pups."[41]
In other species, too, interest in copulation for its own sake signaled the weakness of female character. A late-eighteenth-century agriculturalist criticized the female ass for being "full as lascivious" as the male, which he claimed made her "a bad breeder, ejecting again the seminal fluid she has just received in coition, unless the sensation of pleasure be immediately removed by loading her with blows." Similarly, he condemned the sow, who "may be said to be in heat at all times; and even when she is pregnant she seeks the board, which, among animals, may be deemed an excess." An edifying contrast, he pointed out, was offered by the demure behavior of cows, which, once pregnant, "will not suffer the bull to approach them."[42]
In males, however, eagerness to copulate was matter for praise. The he-goat was admired as "no despicable animal . . . so very vigorous . . . that one will be sufficient for above a hundred and fifty she-goats."[43] Breeders agreed that the only reason to curt the enthusiasm of studs was physical rather than moral, since too-frequent copulation was feared to undermine the constitution of both sire and offspring. Thus, one authority on horses complained that "our test stallion . . . cover too many mares in one season; and this is the reason why they get so few good colts"; another advised against pasturing a stallion with a herd of mares because in this situation "in six weeks, [he] will do himself more damage than in several years by moderate exercise."[44] Similarly, one expert on pedigreed dogs warned, "If you possess a champion dog . . . do not be tempted to stud him too much, or you may kill the goose which lays the eggs of gold. One bitch a fortnight is about as much as any dog can do, to have good stock and retain his constitution."[45]
Despite the need to practice such precise accounting, the sexual management of male animals was much simpler than that of females. In the company of a suitable partner, bulls, stallions, dogs, boars, and rams ordinarily did just what was expected of them. But even after breeders had presented their female animal to a suitable male in the required receptive and unsullied condition, their cares were not over. At that point, the female might decide to exercise an inappropriate veto power, offering an unmistakable challenge to the authority of her owner. After all, in an enterprise dedicated to the production of offspring, too much reluctance was as bad as too little, and resistance of legitimate authority was as unfeminine as proscribed sexual enjoyment.
The terms in which breeder described such insubordination expressed not only the anger it provoked but the extent to which that anger reflected worries about sexual subordination within their own species. Some categories of females were viewed with special suspicison. For exam-
ple, bitches of the larger breeds, whose physical endowments commanded respect whether or not they were feeling refractory, had "to be taped or muzzled . . . to prevent either yourself or the dog from being bitten." Maiden bitches, too, were "generally a great annoyance from first to last." Their coyness might have to be countracted with coercion, although breeders were cautined to remember, "not too much."[46] But almost any kind of bitch might evince reluctance when confronted with a prospective made not of her ow choosig, in which case, she could be castigated as "troublesome," "morose," or even "savage."[47] The prescribed remedy was "hard exercise, until the bitch is thoroughly exhausted"; often this would "reduce a tiresome animal to submission." Bitches that reflused to particpate willingly at this point provoked their owners to severer measures—measures that again recalled the clichés of romantic fiction. They might be "drugged with some narcotic," or, in the most serious cases of insubordination, the male dog and the human breeder might cooperate in what was referred to as a "forced service."[48]
The technical discourse of animal husbandry clearly reflected contemporary concerns about human gender relations, even though the eighteenth- and nineteenth-century breeders of domesticated animals were not explicitly aware of this subtext as they shared their practical wisdom and rehearsed their triumphs. But they would not have made the same claims about the nature of their animals or used the same language to describe bovine and canine inclinations and behavior if they had not implicitly assumed an identity between human nature and animal nature—an identity they would certaily have denied if they had been asked about it directly. From the perspective of the late twentieth century, this particular conflation of the human social world with that of domestic animals is easy to criticize. Their projection of human social constructions onto animal mating habits led breeders astray in some practical senses, introducing inefficiency along with misunderstanding and error. And it expressed a view of gender relations that seems both incorrect and objectionable by contemporary standards. But it is hard not to feel that by placing people among the other animals, breeders also implicitly acknowledged an underlying truth, one that human beings have traditionally striven to deny, no less today than in the past.
Four—
Language and Ideology in Evolutionary Theory:
Reading Cultural Norms into Natural Law
Evelyn Fox Keller
In the mid-twentieth century, biology became a "mature science," that is, it succeeded, finally, in breaking through the formidable barrier of "life" that had heretofore precluded it from fully joining the mechanicoreductive tradition of the physical sciences. For the first time in history, the "secret of life" could credibly be claimed to have been solved: Living beings—presumably including man along with the rest of the animal kingdom—came to be understood as (mere) chemical machines.
But the chemical machine that constitutes a living organism is unlike machines of the eighteenth and nineteenth centuries. It is not a machine capable only of executing the purposes of its maker, that is, man, but a machine endowed with its own purpose. In short, it is a machine of the twentieth century, a cybernetic machine par excellence: absolutely autonomous, capable of constructing itself, maintaining itself, and reproducing itself. As Jacques Monod has explained:
. . . The entire system is totally, intensely conservative, locked into itself, utterly impervious to any "hints" from the outside world. Through its properties, by the microscopic clockwork function that establishes between DNA and protein, as between organism and medium, an entirely one-way relationship, this system obviously defies any "dialectical" description. It is not Hegelian at all, but thoroughly Cartesian: the cell is indeed a machine . (110–111)
The purpose—the sole purpose—of this machine is its own survival and reproduction, or perhaps more accurately put, the survival and reproduction of the DNA programming and "dictating" its operation. In Richard Dawkins's terms, an organism is a "survival machine," a "lumbering robot" constructing to house its genes, those "engines of self-preser-
vation" that have as their primary property that of being inherently "selfish." They are "sealed off from the outside world, communicating with it by tortuous indirect routes, manipulating it by remote control. They are in you and in me; they created us, body and mind; and their preservation is the ultimate rationale for our existence" (21). With this description, man himself has become a machine, but perhaps it might alternatively be said that the machine itself has become man.
The general question is this: To what extent can our contemporary scientific descrption of animate forms, culminating in the description of man as a chemical machine, be said to be strictly technical, and to what extent does it actually encode particular conceptions of man—conceptions that derive not so much from a technical domain as from a social, political, and even psychological domain? Have animate, even human, forms finally been successfully deanimated and mechanized, or have their mechanical representations themselves been inadvertently animated, subtly recast in particular images of man?
I suggest that traces of such images might be found in virtually all scientific representations of nature, but they are perhaps especially conspicuous in our descriptions of the evolution of animate forms—even in those representations that make the greatest claims to conceptual neutrality. It is no secret that evolutionary biology has provided a particularly fertile field for those who seek to demonstrate the impact of social expectations on scientific theory. Indeed, it might be said that it is precisely for this reason that modern evolutionary theorists have sought so strenuously to place their discipline on firm scientific footing. Population genetics and mathematical ecology are the two subdisciplines that have been constructed to meet this need—to provide a rigorous substructure for all of evolutionary biology. The general methodological assumption that underlies both of these subdisciplines can be described as atomic individualism, that is, the assumption that a composite property of a system both can and should be represented by the aggregation of properties inhering in the individual atoms constituting that system, appropriately modified by their pairwise or higher order interactions.[1]
As is conventional in biological discourse, I take the individual atom to be, alternatively, the organism or the gene. I shall, therefore, focus on the particular attributes customarily assumed to characterize the basic unit of analysis, the individual organism or gene.
But my focus will be on the practice rather than the principle of atomic individualism in evolutionary theory. Others hae argued for an ideological load in the very assumptions of this methodological orientation (see, e.g., Wimsatt, Dupré), but here I wish to bracket such questions and focus instead on the lack of neutrality in its actual applications. In particular, I claim that properties of the "individual" that are generally
assumed to be necessary are in fact contingent, drawn not from nature but from our own social and psychosocial heritage. More specifically, I will argue that much of contemporary evolutionary theory relies on a representation of the "individual"—be it the organism or the gene—that is cast in the particular image of man we might call the "Hobbesian man": simultaneously autonomous and oppositional, connected to the world in which it finds itself not by the promise of life and growth but primarily by the threat of death and loss, its first and foremost need being the defense of its boundaries. In psychological terms, we might say that such an individual betrays an idealized conception of autonomy: one that presupposes a radical conception of self and that simultaneously attributes to the relation between self and other an automatic negative valence, a relation, finally, not so much of independence as of dynamic opposition.
I claim that this psychosocial load is carried into evolutionary theory not by explicit intention but by language—by tacit linguistic conventions that privilege the autonomy of the individual at the expense of biologically constitutive interactions and, at the same time, obscure the logical distinction between autonomy and opposition. In this, they support the characterization of the biological individual as somehow "intrinsically" competitive, as if autonomy and competition were semantically equivalent, collapsed into one by that fundamentally ambiguous concept, self-interest. Accordingly, it is the languageof autonomy and opposition in contemporary evolutionary theory that is the specific object of my concern.
Discourse of Self and Other
I begin with a relatively accessible example of actual confusion between autonomy and opposition that is found not in the theoretical literature per se but in a more general genre of scientific discourse, at once popularizing and prescriptive. Here the focus is not so much on the independence of one individual from another, of self from other, as on the independence of the most abstract other from self—of nature from man. Accordingly, the negative value that tacitly accrues to this relation attaches not so much to the self as to the other, that is, to nature.
With Darwin, evolutionary biology joined a tradition already well established in the physical sciences—a tradition that teaches that the laws of nature are, in Steven Weinberg's words, "as impersonal and free of human values as the rules of arithmetic" (Weinberg 1974). But this rhetoric goes beyond impersonality: nature becomes uncaring and "hostile," traits that are impersonal in a quite personal sense. To illustrate this tendency, consider, for example, Weinberg's own elaboration of his message:
It is almost irresisible for humans to believe that we have some special relation to the universe, tht human life is not just a more-or-less farcical outcome of a chain of accidents reaching back to the first three minutes. . . . It is very hard to realize that this all is just a tiny part of an overwhelmingly hostile universe. (Midgley, 88)
In much the same vein, Jacques Monod writes,
If he accepts this message in its full significance, man must at last wake out of his millenary dream and discover his total solitude, his fundamental isolation, he must realize that, like a gypsy, he lives on the boundary of an alien world, a world that is deaf to his muisc, and as indifferent to his hopes as it is to his suffering or his crimes.[2] (Monod, 2)
The world we must steel ourselves to accept is a world of "uncaring emptiness," a "frozen universe of solitude" (173). The natural world from which animism has been so carefully expunged has become not quite neutral but "empty," "frozen," "overwhelmingly hostile," and "terrifying."
For the record, though, it was a poet, not a scientist, who gave us our most familiar metaphor conflating an absence of benevolence in nature with "overwhelming hostility." A nature that does not care for us becomes indeed a nature "red in tooth and claw"—callous, brutal, even murderous. It is a nature that cries, "I care for nothing; all shall go."
Mary Midgley suggests that such residual animism properly belongs to what she calls "the drama of parental callousness":
First, there is the tone of personal aggrievement and disillusion, which seems to depend . . . on failure to get rid of the animism or personficiation which (these scientists) officially denounce. An inanimate universe cannot be hostile. . . . Only in a real, conscious human parent could uncaringness equal hostility. . . . Certainly if we expect the non-human world around us to respond to us as a friendly human would, we shall be disappointed. But this does not put it in the position of a callously indifferent human.[3] (1985: 87)
Midgley's explanation is persuasive—perhaps precisely because the slippage between an indifferent and a hostile nature so clearly does denote a logical error, once pointed out. But a similar problems surfaces in another set of contexts as well—where the move from a neutral to a negative valence in the conceptualization of self-other relations is less evidently a simple "mistake." Here it is not nature but the individual organism, the firs rather than the second term of the self-other dichotomy, whom the insistently unsentimental biologist taxes with hostility.
I am referring in particular to the tradition among evolutionary biologists that not only privileges the individual descriptively but that also, in the attempt to locate all relevant causal dynamics in the properties intrin-
sci to the individual, tends to attribute to that individual not simply autonomy but an additional "intrinsic" competitive bent, as if independence and competition were inseparable traits. The very same move that defines self-interest and altruism as logically opposed makes independence virtually indistinguishable from competition. Michael Ghiselin is one of the most extreme representatives of this position and provides some particularly blatant examples of the rhetorical (and conceptual) conflation I am speaking of. To dramatize his position, he concludes:
The economy of nature is competitive from beginning to end. . . . No hint of genuine charity ameliorates our vision of society, once sentimentatlism has been laid aside. . . . Given a full chance to act for his own interest, nothing but expediency will restrain [an organism] from brutalizing, from maiming, from murdering—his brother, his mate, his parent, or his child. Scratch an "altruist" and watch a "hypocrite" bleed. (247)
Of course, Ghiselin's language is intended to shock us—but only to underscore his thesis. In this effort, he is counting on our acceptance, as readers, first, of the rule of self-interest as logically equivalent to the absence of altruism or charity, and second, of competitive exploitation as a necessary concomitant of self-interest. Our usual willingness to accept these assumptions, or rather, to allow them to pass unnoticed, is itself a measure of the inaccessibility of a domain where self-interest and charity (or altruism) conjoin and, correlatively, of a distinction between self-interest and competition. Unlike the previous example, where no one, if pressed, would say that nature "really is" hostile, Ghiselin's assumptions do seem to accord with the way things "really are." Because the difference between self-interest and competition is less obvious to most of us than the difference between impersonality in nature and hostility, the problem here is much more difficult. In other words, before we can invoke a psychosocial explanation of the conceptual conflation between radical individualism and competition, we need first to see them as different.
We can do this best by turning our attention from a prescriptive discourse that aims to set ground rules for evolutionary theory to an examination of uses of the same language in the actual working out of particular theoretical strategies. I want to look, therefore, at the technical uses of the language of individualism in mathematical ecology and population genetics—in the first case, on the language of competition, and, in the second, on the language of reproductive autonomy. In particular, I want to show how certain conventional interchanges, or trade-offs, between to show how certain conventional interchanges, or trade-offs, between technical and colloquial language cast a blanket of invisibility, or rather, of unspeakability, over certain distinctions, categories, and questions. It is, I suggest, precisely through the maintenance of such an aura of unspeakability that social, psychological, and political expectations
generally exert their influence, through language, on the actual structure and content of scientific theory.
Competition and Cooperation in Mathematical Ecology
One problem I want to examine arises in the systematic neglect of cooperative (or mutulaist) interactions and the correspondig privileging of competitive interactions evident throughout almost the entire history of mathematical ecology. When we ask practitioners in the field for an explanation of this historical disinterest in mutualist interactions, their response is usually one of puzzlement—not so much over the phenomenon as over the question. How else could it, realistically, be? Yes, of course, mutualist interactions occur in nature, but they are not only rare, they are necessarily secondary.
Often it is assumed that they are in the service of competition: such pheomena have at times actually been called "cooperative competition." Indeed, the expectation of most workers in the field that competition is both phenomenologically primary and logically prior is so deeply embedded that the very question has difficulty getting airspace: there is no place to put it. My question thus becomes, What are the factors responsible for the closing-off of that space?
Part of the difficulty in answering this question undoubtedly stems from the massive linguistic confusion in conventional use of the term competition. One central factor can be readily identified, however, and that is the recognition that, in the real world, resources are finite and hence ultimately scarce. To most minds, scarcity automatically implies competition, both in the sense of "causing" competitive behavior and in the sense of constituting, in itself, a kind of de facto competitio, independent of actual interactions between organisms. Indeed, so automatic is the association between scarcity and competition that, in modern ecological usage, competition has come to be defined as the simultaneous reliance of two individuals, or two species, on an essential resource that is in limited supply (see, e.g., Mayr 1963: 43). Since the scarcity of resources can itself hardly be questioned, such a definition lends to competition the same a priori status.
This technical definition of competition was probably first employed by Voltera, Lotka, and Gause in their early attempts to provide a mathematical representatation of the effects of scarcity on the population growth of "interacting" species, but it soon came to be embraced by a wider community of evolutionary biologists and ecologists—partly, at least, to neutralize the discourse and so bypass the charge of ideologically laden expectations about (usually animal) behavior, in fact freeing the discourse of any depen-
dence on how organisms actually behave in the face of scarcity. The term competition now covered apparently pacific behavior just as well as aggressive behavior, an absurdity in ordinary usage but protected by the stipulation of a technical meaning. As Ernst Mayr explains,
To certain authors eve since [Darwin], competition has meant physical combat, and, coversely, the absence of physical combat has been taken as an indication of the absence of competition. Such a view is erroneous. . . . The relatively rarity of overt manifestations of competition is proof not of the insignificance of competition, as asserted by some authors, but, on the contrary, of the high premium natural selection pays for the development of habits or preferences that reduce the severity of competition. (1963: 42–43)
Paul Colinvaux goes one step farther, suggesting that "peaceful coexistence" provides a better description than any "talk of struggles for survival." "Natural selection designs different kinds of animals and plants so that the avoid competition. A fit animal is not one that fights well, but one that avoids fighting altogether" (1978: 144).
But how neutral in practice is the ostensibly technical use of competition employed both by Mayr and Colinvaux? I suggest two ways in which, rather than bypassing ideological expectations, it actually preserves them, albeit in a less visible form—a form in which they enjoy effective immunity from criticism. so as not to be caught in the very trap I want to expose, let me henceforth denote competition in the technical sense as "Competition" and in the colloquial sense (of actual contest) as "competition."
The first way is relatively straightforward. The use of a term with established colloquial meaning in a technical context permits the simultaneous transfer and denial of its colloquial connotations. Let me offer just one example: Colinvaux's own description of Gause's original experiments that were designed to study the effect of scarcity on interspecific dynamics—historically, the experimental underpinning of the "competitive exclusion coexistence." He writes,
No matter how many times Gause tested [the paramecia] against each other, the outcome was always the same, complete extermination of one species. . . . Gause could see this deadly struggle going on before his eyes day after day and always with the same outcome. . . . what we [might have] expected to be a permanent struggling balance in fact became a pogrom. (142)
Just to set the record straight, these are not "killer" paramecia but perfectly ordinary paramecia—minding their own business, eating and dividing, or not, perhaps even starving. The terms extermination, deadly struggle , and program refer merely to the simultaneous dependence of two species on a common resource. If, by chance, you should have misinter-
preted and taken them literally, to refer to overt combat, you would be told that you had missed the point. The Lotka-Volterra equations make no such claims; strictly speaking, they are incompatible with an assumption of overt combat; the competitive exclusion principle merely implies an avoidance of conflict. And yet the description of such a situation, only competitive in the technical sense, slips smoothly from "Competition" to genocide—much as we saw our neo-Tennysonians slip from inpersonality to heartless rejection.
The point of this example is not to single out Colinvaux (which would surely be unfair) but to provide an illustration of what is, in fact, a rather widespread investment of an ostensibly neutral technical term with a quite different set of connotations associated with its colloquial meaning. The colloquial connotations lead plausibly to one set of inferences and close off others, while the technical meaning stands ready to disclaim responsibility if challenged.[4]
The second and more serious route by which the apparently a priori status of competition is secured can be explored through an inquiry into the implicit assumptions about resource consumption that are here presupposed and the aspects of resource consumptions that are excluded. The first presupposition is that a resource can be defined and quantitatively assessed independent of the organism itself; and the second is that each organism's utilization of this resource is independent of other organisms. In short, resource consumption is here represented as a zerosum game. Such a representation might be said to correspond to the absolutely minimal constraint possible on the autonomy of each individual, but it is a constraint that has precisely the effect of establishing a necessary link between self-interest and competition. With these assumptions, apparently autonomous individuals are in fact bound by a zerosum daynamic that guarantees not quite an absence of interaction but the inevitability of a purely competitive interaction. In a world in which one organism's dinner necessarily means another's starvation, the mere consumption of resources has a kind of de facto equivalence to murder: individual organisms are locked into a life and death struggle not by virtue of their direct interactions but merely by virture of their existence in the same place and time.
It is worth noting that the very same (Lotka-Volterra) equations readily accommodate the replacement of competitive interactions by cooperative ones and even yield a stable solution. This fact was actually noted by Gause himself as early as 1935 (Gause and Witt 1935) and has been occasionally rediscovered since then, only to be, each time, reforgotten by the community of mathematical ecologists. The full reasons for such amnesia are unclear, but it suggests a strong prior commitment to the representation of resource consumption as a zero-sum dynamic—a
representation that would be fatally undermined by the substitution (or even addition) of cooperative interactions.
Left out of this representation are not only cooperative interactions but any interactions between organisms that affect the individual's need and utilization of resources. Also omitted are all those interactions between organism and environment which interfere with the identification and measurement of a resource independently of the properties of the organism. Richard Lewontin (1982) has argued that organisms "determine what is relevant" in their environment—what, for example, is a resource—and actually "construct" their environment. But such interations, either between organisms or between organism and environment, lead to pay-off matrices necessarily more complex than those prescribed by a zero-sum dynamic—pay-off matrices that, inturn, considerably complicate the presumed relation between self-interest and competition, if they do not altogether undermine the very meaning of self-interest.
Perhaps the simplest example is provided by the "prisoner's dilemma." But even here, where the original meaning of self-interest is most closely preserved, Robert Axelrod has shown that under conditions of indefinite reiterations, a ("tit-for-tat") strategy is generally better suited to self-interest than are more primitive competitive strategies.
Interactions that effectively generate new resources, or either increase the efficiency of resource utilization or reduce absolute requirement, are more directly damaging to the very principle of self-interest. These are exactly the kinds of interactions that are generally categorized as special cases: as "mutualist," "cooperative," or "symobiotic" interactions. Finally, interactions that affect the birth rate in ways not mediated by scarcity of resources, for example, sexual reproduction, are also excluded by this representation. Perhaps the most important of these omissions for intraspecific dynamics, I would point to sexual reproduction, a fact of life that potentially undermines the core assumptions of radical individualism. In the last few years, there has been a new wave of interest in mutualism among not only dissident but even a few mainstream biologists, and numerous authors are hard at work redressing the neglect of previous years.[5] But in the sixty years in which the Lotka-Volterra equations have reigned as the principal, if not the only, model of interspecific population dynamics—even in the more genial climate of recent years—the omission of sexual reproduction from this model has sacrcely been noted.
This omission, once recognized, takes us beyond the question of selective biases in admissible or relevant interactions between organisms. It calls into question the first and most basic assumption for the methodology of individualism in evolutionary theory, namely, that in-
trinsic properties of individual organisms are primary to any description of evolutionary phenomena.[6] To examine this argument, let us turn from mathematical ecology to population genetics, that branch of evolutionary theory that promises to avoid the practical difficulties of selective focus on certain interactions by excluding the entire question of competitive or cooperative interactions from its domain. In other workds, traditional population genetics addresses neither interactions betwee organisms nor limitations in resources; it effectively assumes populations at low density with infinite resources.
However, one last problem with the language of competition must be noted lest it carry over int our discussion of individual autonomy in population genetics: the widespread tendency to extend the sense of "competition" to include not only the two situations we distinguished earlier (i.e., conflict and reliance on a common resource) but also a third situation[7] where there is no interaction at all, where "competition" denotes an operation of comparison between organisms (or species) that requires no juxtaposition in nature, only in the biologist's own mind. This extention, where "competition" can cover all possible circumstances of relative viability and reproductivity, brings with it, then, the tendency to equate competition with natural selection itself.
Darwin's own rhetorical equation between natural selection and the Malthusian struggle for existence surely bears some responsibility for this tendency. But contemporary readers of Darwin like to point out that he did try to correct the misreading his rhetoric invited by explaining that he meant the term struggle in "a large and metaphoric sense," including, for example, that of the plant on the edge of the desert: competition was only one of the many meanings of struggle for Darwin. Others have been even more explicit on this issue, repeatedly noting the importance of distinguishing natural selection from "a Malthusian dynamic." Lewontin, for one, has written.
Thus, although Darwin came to the idea of natural selection from consideration of Malthus' essay on overpopulation, the element of competition between organisms for a resource in short supply is not integral to the argument. Natural selection occurs even when two bacterial strains are growing logarithmcally in an excess of nutrient broth if they have different division times. (1970: 1)
However, such attempts—by Lewontin, and earlier and more comprehensively, by L.C. Birch (1957)—to clarify the distinction between natural section and competition (what Engels called "Darwin's mistake") have done little to stem the underlying conviction that the two are somehow the same. In a recent attempt to define the logical essence of "the Darwinian dynamic," Bernstein et al. (1983) freely translate Darwin's
"struggle for survival" to "competition through resource limitation" (192), thereby claiming for competition the status of a "basic component" of natural selection. Even more recently, George Williams (1986) describes a classic example of natural selection in the laboratory as a "competition experiment," a "contest" between a mutant and normal allele, in which he cites differential fecundity as an example of "the competitive interactions among individual organisms" that cause the relative increase in one population (114–115).
At issue is not whether overtly competitive behavior or more basic ecological scarcity is the rule in the natural world; rather, it is whether or not such a question can even be asked. To the extent that distinctions between competition and scarcity, on the one hand, and between scarcity and natural selection, on the other, are obliterated from our language and thought, the question itself becomes foreclosed. As long as the theory of natural selection is understood as a theory of competition, confirmation of one is taken to be confirmation of the other, despite their logical (and biological) difference.
While this clearly raises problems about the meaning of confirmation, my principal concern is with the dynamics by which such an oversight or confusion is sustained in the theory and practice of working biologists—with the internal conventions that render it effectively resistant to correction. Dynamics similar to those in the language of competition can also be seen in the language of reproductive autonomy, especially as employed in the theory and practice of population biology.
The Problem of Sexual Reporoduction
In much of the discourse on reproduction, it is common to speak of the "reproduction of an organism"—as if reproduction is something an individual organism does; as if an organism makes copies of itself, by itself. Strictly speaking, of course, such language is appropriate only to asexually reproducing populations since, as every biologist knows, sexually reproducing organisms neither produce copies of themselves nor produce other organisms by themselves. It is a striking fact, however, that the language of individual reproduction, including such correlative terms as an individual's offspring and lineage , is used throughout population biology[8] to apply indiscriminately to both sexually and asexually reproducing populations. While it would be absurd to suggest hat users of such language are actually confused about the nature of reproduction in the organisms they study (e.g., calculations of numbers of offspring per organism are always appropriately adjusted to take the mode of reproduction into account), we might nonetheless ask, what functions, both positive and negative, does such manifestly peculiar language
serve? And what consequences does it have for the shape of the theory in which it is embedded?
I want to suggest, first, that this language, far from being inconsequential, provides crucial conceptual support for the individualist program in evolutionary theory. In particular, my claim is that the starting assumption of this program—that is, that individual properties are primary—depends on the language of individual reproduction for its basic credibility.[9] In addition, I would argue that, just as we saw with the language of competition, the language of individual reproduction, maintained as it is by certain methodological conventions, both blocks the perception of problems in the evolutionary project as presently conducted and, simultaneously, impedes efforts to redress those difficulties that can be identified.
The problems posed for evolutionary theory by sexual reproduction and Mendelian genetics are hardly new, and indeed, the basic theory of population genetics originates in the formulation of a particular method (i.e., the Hardy-Weinberg calculus) designed to solve these problems. The Hardy-Weinberg calculus (often referred to as "bean-bag" genetics) invoked an ibviously highly idealized representation of the relation between genes, organisms, and reproduction, but it was one that accomplished a great deal. Most important, it provided a remarkably simple recipe for mediating between individuals and populations—a recipe that apparently succeeded in preserving the individualist focus of the evolutionists' program. One might even say that it did so, perhaps somewhat paradoxically, by tacitly discounting individual organisms and their troublesome mode of reproduction. With the shift of attention from populations of organisms to well-mixed, effectively infinite, pools of genes, the gap between individual and population closed. Individual organisms, in this picture, could be thought of as mere bags of genes (anticipating Richard Dawkins's "survival machines" [1976: 21])—the end product of a reproductive process now reduced to genetic replication plus the random mating of gametes. Effectively bypassed with this representation were all the problems entailed by sexual difference, by the contingencies of mating and fertilization that resulted from the finitude of actual populations and, simultaneously, all the ambiguities of the term reproduction as applied to organisms that neither make copies of themselves nor reproduce by themselves. In shor, the Hardy-Weinberg calculus provided a recipt for dealing with reproduction that left undistrubed—indeed, finally, reinforced—the temptation to think (and to speak) about reproduction as simply an individual process, to the extent, that is, that it was thought or spoken about at all.
In the subsequent incorporation of the effects of natural selection into the Hardy-Weinberg model, for most authors in population genetics, the
contribution of reproduction to natural selection fell largely by the wayside. True, the basic calculus provided a ready way to incorporate at least part of the reproductive process, namely, the production of gametes; but in practice, the theorietical (and verbal) convention that came to prevail in traditional population genetics was to equate nautral selection with differential survival and ignore fertility altogether. In other words, the Hardy-Weinberg calculus seems to ave invited not one but two kinds of elision from natural selection—first, of all those complications incurred by sex and the contingency of mating (these, if considered at all, bet shunted off under the label of sexual, rather than natural, selection),[10] and second, more obliquely, of reporudction in toto.
In want to suggest that these two different kinds of elision in fact provided important tacit support for each other. In the first case, the representation of reproduction as gametic production invited confidence in the assumption that, for calculating changes in gene frequency, differential reproduction, or fertility, was like differential survival and hence did not require separate treatment. And in the second case, the technical equation of natural selection with differential survival which prevailed for so many years, in turn, served to deflect attention away from the substantive difficulties invoked in representing reproduction as an individual process. The net effect has been to establish a circle to confidence, first, in the adequacy of the assumption that, despite the mechanics of Mendelianism, the individual remains both the subject and object of reproduction, and second, in the adequacy of the metonymic collapse of reproduction and survival in discussions of natural selection.
The more obvious cost of this circle surely comes from its second part. As a number of authors have recently begun to remind us, the equation between natural selection and differential survival fosters both the theoretical omission and the experimental neglect of a crucial component of natural selection. Perhaps even more serious is the cost in unresolved difficulties that this equation has helped obscure.
One such difficulty is the persistence of a chronic confusion between two definitions of individual fitness: one, the (average) net contribution of an individual of a particular genotype to the next generation, and the other, the geometric rate of increase of that particular genotype. The first refers to the contribution an individual makes to reproduction, while the second refers to the rate of production of individuals. In other words, the first definition refers to the role of the individual as subject of reproduction and the second to its role as object. The disparity between the two derives from the basic fact that, for sexually reproducing organisms, the rate at which individuals of a particular genotype are born is a fundamentally different quantity from the rate at which individuals of
that genotype give birth—a distinction easily lost in a language that assigns that same term, birthrate , to both processes.
Beginning in 1962, a number of authors have attempted to call attention to this confusion (Moran 1962; Charlewsorth 1970; Pollak and Kempthorne 1971; Denniston 1978), agreeing that one definition—the contribution a particular genotype makes to the next generation's population—is both conventional and correct, while the other (the rate at which individuals of a particular genotype are born) is not. Despite their efforts, however, the confusion persists.[11] In part, this is because there remains a real question as to what "correct" means in this context or more precisely, as to which definition is better suited to the needs that the concept of fitness is intended to serve—in particular, the need to explain changes in the genotypic composition of populations. Given that need, we want to know not only which genotypes produce more but also the relative rate of increase of a particular genotype over the course of generations.
Not surprisingly, conflation of the two definitions of fitness is particularly likely to occur in attempts to establish a formal connection between the models of population genetics and those of mathematical ecology. Because all the standards models for population growth assume asexual reproduction, the two formalisms actually refer to two completely different kinds of populations: one of gametic pools and the other of asexually reproducing organisms. In attempting to reconcile these two theories, such a conflation is in fact require to finesse the logical gap between them. A more adequate reconciliation of the two formalisms requires the introduction of both the dynamics of sexual reproduction into mathematical ecology and a compatible representation of those dynamics into population genetics.
Counterintuitively, it is probably the second—the inclusion (in population genetics models) of fertility as a property of the mating type—that calls for the more substantive conceptual shifts. Over the last twenty years, we have witnessed the emergence of a considerable literature devoted to the analysis of fertility selection—eading at least some authors to the conclusion that "the classical concept of individual fitness is insufficient to account for the action of natural selection" (Christiansen 1983: 75).
The basic point is that when fertility selection is included in natural selection, the fitness of a genotype, like the fitness of a gene (as argued by Sober and Lewontin 1982), is seen to depend on the context in which it finds itself. Now, however, the context is one determined by the genotype of the mating partner rather than by the complementary allele. A casual reading of the literature on fertility selection might suggest that the mating pair would be a more appropriate unit of selection than the
individual, but the fact is that mating pairs do not reproduce themselves any more than do individual genotypes. As E. Pollak has pointed out, "even if a superior mating produces offspring with a potential for entering a superior mating, the realization of this potential is dependent upon the structure of the population" (1978: 389). In other words, in computing the contribution of either a genotype or a mating pair to the next generation's population (of genotypes or mating pairs), it is necessary to take account of the contingency of mating: such a factor, measuring the probability that any particular organism will actually mate, incurs a frequency dependence that reflects the dependence of mating on the genotypic composition of the entire population.
Very briefly, the inclusion of a full account of reproduction in evolutionary theory necessitates the conclusion that natural selection operates simultaneously on many levels (gene, organisms, mating pair, and group), not just under special circumstances, as others have argued, but as a rule. For sexually reproducing organisms, fitness in general is not an individual property but a composite of the entire interbreeding population, including, but certainly not determined by, genic, genotypic, and mating pair contributions. By undermining the reproductive autonomy of the individual organism, the advent of sex undermines the possibility of locating the causal efficacy of evolutionary change in individual properties. At least part of the "causal engine" of natural selection must be seen as distributed throughout the entire population of interbreeding organisms.
My point is not merely to argue against the adequacy of the individualist program in evolutionary theory but—like the point of my earlier remarks about competition—to illustrate a quiet general process by which the particular conventions of language employed by a scientific community permit a tacit incorporation of ideology into scientific theory and, simultaneously, protect participants from recognition of such ideological influences. The net effect is to insulate the theoretical structure from substantive critical revision. In discussions of sexual reproduction, the linguistic conventions of individual reproduction—conventions embodying an ideological commitment to the a priori autonomy of the individual—both perpetuate that belief and promte its incorporation into the theory of evolutionary dynamics. At the same time, the conventional equation between natural selection and differential survival has served to protect evolutionary theory from problems introduced by sexual reproduction, thereby lending at least tacit support to the assumption of individual autonomy that gave rise to the language of individual reproduction in the first place. The result—now both of the language of autonomy and the language of competition—is to effectively exclude from the domain of theory those biological phenomena that do not fit (or even worse, threaten to undermine) the ideological commitments
that are unspoken yet in language, built into science by the language we use in both constructing and applying our theories. In this way, through our inescapable reliance on language, even the most ardent efforts to rid natural law of cultural norms become subverted, and the machinery of life takes on not so much a life of its own as a life of ur own. But then again, what other life could it have?
References
Axelrod, Robert. The Evolution of Cooperation. New York: Basic Books, 1984.
Bernstein, H., H. C. Byerly, F. A. Hopf, R. A. Michod, and G. K. Vemulapalli. "The Darwinian Dynamic," The Quarterly Review of Biology 58 (1983): 185-207.
Birch, L. C. "Meanings of Competition." American naturalist 91 (1957): 5-18.
Boucher, Douglas. The Biology of Mutualism. Cambridge: Oxford University Press, 1985.
Charlesworth, B. "Selection in Populations with Overlapping Generations. 1: The Use of Malthusian Parameters in Population Genetics," Theoretical Population Biology 1, 3 (1970): 352-370.
Christiansen, F.B. "The Definition and Measurement of Fitness," in B. Shorrocks, ed., Evolutionary Ecology: B. E. S. Symposium 23. Oxford and Boston: Blackwell Scientific Publications, 1984. 65-79.
Colinvaux, Paul. Why Big Fiere Animals are Rare. Princeton: Princeton University Press, 1978.
Dawkins, Richard. The Selfish Gene. Oxford University Press, 1976.
Denniston, C. "An Incorrect Definition of Fitness Revisited," Annals of Human Genetics, Lond. 42 (1978): 77-85.
Gause, G. F., and A. A.Witt. "Behavior of Mixed Populations and the Problem of Natural Selection," American Naturalist 69 (1935): 596-609.
Ghiselin, Michael. The Economy of Nature and the Evolution of Sex. Berkeley, Los Angeles, London: University of California Press, 1974.
Keller, Evelyn Fox. "Reproduction and the Central Project of Evolutionary Theory," Biology and Philosophy 2 (1987): 73-86.
Keller, Evelyn Fox. "Demarcating Public from Private Values in Evolutionary Discourse," Journal of the History of Biology 21, 2 (1988): 195-211.
Lewontin, Richard, "Organism and Environment," in E. H. C. Pltokin, ed., Learning, Development, and Culture. New York: John Wiley and Sons, 1982.
———. "The Units of Selection," Annual Review of Ecology and Systematics (1970): 1-1B.
Mayr, Ernst. Animal Species and Evolution. Cambridge: Harvard University Press, 1963.
Midgley, Mary. Evolution as a Religion. London and New York: Methuen, 1985.
Monod, Jacques. Chance and Necessity. New York: Random House, 1972.
Moran. P. A. P. "On the Nonexistence of Adaptive Topographies," annals of Human Genetics , 27 (1962): 383-393.
Pollak, E. "With Selection for Fecundity the Mean Fitness Does Not Necessarily Increase," Genetics 90 (1978): 383-389.
Pollak, E., and O. Kempthorne. "Malthusian Parameters in Genetic Populations, II. Random Mating Populations in Infinite Habitats," Theoretical Population Biology 2 (1971): 357-390.
Sober, E., and R. Lewontin. "Artifact, Cause, and Genetic Selection," Philosophy of Science 47 (1982): 157-180.
Tennyson, Alfred Lord. In Memoriam LV-LVI.
Weinberg, Steven. "Reflections of a Working Scientist," Daedalus 103 (Summer 1974): 33-46.
Williams, George. "Comments," Biology and Philosophy 1, 1 (1986): 114-122.
Five—
Human Nature and Culture:
Biology and the Residue of Uniqueness
Melvin Konner
I
Since the question, "Is there such a thing as human nature?" continues to be posed, and to be answered by some in the negative, it seems necessary to being by exploring the meaning of this question and showing why it cannot, in any meaningful form, have any but an affirmative answer. The question raises doubts about whether aspects of the human condition can be considered inevitable outcomes of biological elements in the human makeup. In extreme forms, such doubts are dubious. If I assert that it is human nature to require a diet supplying eight essential amino acids, to walk bipedally, and to speak a language containing nouns and verbs, there are only a few ways to question the validity of the claim.
For example, it is possible to point to a small number of individuals who, from birth onward, are poisoned by an amino acid essential to all others, or who are never able to walk or speack, but these observations—which might be called the "rare-defect objection"—do not detract from the power of the generalizations. Second, and somewhat more interesting, it is easy to show that great variability exists in the way these generalizations are fulfilled by individuals and groups and that most of this variability is due to cultural or environmental causes. But these observations, the "cross-cultural objection," do not address the point that there is, in each of the systems in question, a core of features that do not vary. Third, there is the "freedom-of-will" objections: some individuals perfectly capable of walking will choose never to do so, while others may learn to walk faster or more gracefully than anyone in the world, or even to walk on their hands.
There are several answers to this objection, but two are decisive. First,
such freedom of will exists, but the rarity with which it is exercised, at least at the extremes of the distribution of behavior, is not irrelevant to the question at hand. This answer resembles the reply to the rare-defect objection but is directed to motivation rather than capability, and it implies that motivations as well as capabilities may be legitimately included in the definitional sphere of human nature, a point to which we will return.
Second, within the definable but admittedly broad band of variation in patterns of walking resulting fromt he exercise of freedom of will, it is possible to show lawful relations between human choices and physiological outcomes. After the choice has been exercised—for a given daily amount of walking, say—there follow predictable consequences for the structure and function of muscle, bone, and the cardiopulmonary organs. These, in turn, have consequences for behavioral capability, health, and life span. The laws relating the precise specturm of choices to the predictable outcomes are also part of what is meant by human nature. (Here perhaps a counterobjection will be raised: "But that is just what I mean. Human behavior is flexible, so much so that biology itself is under the sway of human choices." Of course it is; but predictably so, and with thresholds and limits provided by human nature—by the genes—and not by human choices. Biological concepts such as facultative adaptation, range of reaction, and bounded learning, among others, have been developed to describe parallel phenomena in nonhuman species.)
Finally, the "so-waht objection" holds that the sorts of characteristics legitimately subsumed under the rubric of human nature are trivial and uninteresting. "Of course everybody can walk; so what? It's the variations that are interesting." It might be pointed out that every person who can walk exhibits a relatively stereotyped heel-toe progression with synchronized alternate arm swinging, reflecting a very complex wired-in coordinating pattern involving many millions of neurons. Or that every child assumes the ability to walk within a narrowly defined age period because of the maturation of certain parts of the nervous system, the growth of which are under genetic control. But then each successive complexity pointed out becomes subsumed under the ho-hum or sowhat rubric, so that as soon as it is shown to be biologically determined, it will be deemed uninteresting. "That's not what I mean by human nature" will retreat to encompass only that which has not yet been given a biological basis.
Of course, what most of us mean by human nature is not primarily the dependence on certain nutrients or the ability to walk and talk but the condition of being subject to certain patterns of motivation that seem to constrain human freedom, and to this extent, the so-what objection really does apply to the characteristics we have considered so
far. I will insist int he end that human nature fairly includes, in fact must include, certain characteristically human—that is, genetically coded, wired-in—perceptions, cognitions, competencies, response systems, and motor action patterns as well as motivations. Indeed, it is doubtful whether motivation can meaningfully be defined so as to exclude such other psychological characteristics. But it is necessary at this point for us to leave the safe realms of amino acids and coordinated locomotion for the more treacherous realms of love, fear, sacrifice, selfishness, lust, and violence, which come most readily to the mind when the concept of human nature is invoked, and to establish the extent to which these realms are subject to the kinds of arguments applicable in the safer ones.
To do this properly requires a series of intellectual strategies directed at exploring the phylogenetic origins of the pattern; the evolutionary, principally selective forces that have operated and continue to operate on it; the cross-cultura and other variability in it (or lack thereof) and the lawful relationships between the variability and variations in the environment that might be expected to influence it; the immediate physiological antecedents of the pattern; the sequence of development of the pattern, including both the maturational and environmental components; and the genetics of the pattern, as explored through both classical Mendelian methods and the methods of molecular and developmental genetics.
Each of these intellectual strategies has progressed so far in the last two decades as to render suspect any opinion on human nature that does not give clear evidence of having kept abreast of their progress. It is not possible here to review their implications even in relation to one of the motivational patterns of interest. However, it is possible to touch on some of the highlights relevant both to human motivation in general and to a legitimate concept of human nature (Konner 1982b ).
II
The fossil evidence for human and proto-human evolution has accumulated steadily for more than one hundred years so no one familiar with it doubts the reality and continuity of that evolution (Ciochon and Fleagle 1987), However, new discoveries are made each year which change the details of the picture, and during the past tow decades, biochemical taxonomy has further altered our concept of human evolution. There is, therefore, little to be gained for our purposes in closely following each argument in human paleontology. But there is much to be gained from understanding the general higher primate background of human evolution; the environment of human evolutionary adaptedness, that of hunt-
ing and gathering; and the principles of evolutionary adaptation as applied to behavior and reproduction.
All higher primates without exception are social animals with great learning capacity and with the mother-offspring bond at the center of social life (Cheney, Seyfarth, and Smuts 1986). This bond is always prolonged, as is the anatomic and behavioral course of individual development, including each phase of the life cycle as well as the life span as a whole. Laboratory and field studies alike demonstrate the capacity for complex social cognition and social learning, up to and including "cultural" transmission of social rank, tool-using techniques, and knowledge of location of food sources. Play, especially social play, is characteristic of all primate species, particularly during development, and there are various reasons to believe it to be an important opportunity for learning. For example (MacLean 1985), the higher primate emphasis on both the mother-infant bond and on play represents an intensification of the pattern established by the early mammals and is essential to the understanding of the phylogeny of the limbic system and the emotions.
Primate groups generally include a core of genetically related individuals with associated nonrealtives. In most instances, the core is a matrilineage, stable over the life course of the individuals. In any case, the distribution of acts of social support and generosity is preferentially toward genetic relatives but not exclusively so. Monkeys and apes aid nonrelatives and can usually expect some aid in return. Cooperation is ubiquitous, but so is competition, and one of the major purposes of cooperation is mutual defense against other group members. Conflict is frequent, with both sexes participating but with males generally exhibiting more physical aggression than females.
Beyond these broad generalizations, great variation exists in social organization both between and within species (Napier and Napier 1985). Monogamy is present in some South American monkeys, in gibbons, and in one or two other forms, but in most species, larger group associations subsuming more temporary (although sometimes more than transient) associations between individual males and individual females are the rule. The causes of this variation in higher primate social organization remain obscure, but some relevant evolutionary principles will be considered below.
Although there are important species variations, it may be generally said that higher primates are sensitive to significant perturbations of the early social environment, such as isolation rearing or repeated involuntary mother-infant separation, and that these perturbations give rise to abnormalities of sexual, maternal, and aggressive behavior that in humans would be viewed as psychopathology (Harlow 1963, 1969; Arling and Harlow 1967; Sackett et al. 1981). In a number of species
(though not all), isolation rearing gives rise to stereotyped behavior such as rocking and self-directed aggression, and mother-infant separation gives rise to symptoms usually described as protest followed by depression. Even deprivation of contact with peers during development has produced abnormal behavior in some experiments. Apparent human analogues of these causal relationships, although difficult to interpret, have encouraged the use of primate models. These experiments emphasize the extent to which the noraml development and behavior in such animals has come to depend, in the course of evolution, on an intact social matrix.
Natural variation in stable individual behavior patterns ("personality") occurs in free-ranging monkey and ape groups (Smuts 1985) and extends to variants that would be considered pathological if seen in humans, such as hyperaggressive, isolative, phobic, or depressed behavior. It is rarely possible to explore the causes of such variants, but most are probably both genetic and environmental in origin. Some abnormalities, such as severe depression (as in the case of an eight-year-old wild chimpanzee after the death of his mother) may be incompatible with survival (Goodall 1986). Others, however, such as hyperaggressiveness (as in the case of two female chimpanzees that systematically and repeatedly killed the infants of other females) may actually enhance reproductive adaptation for the abnormal individual.
The above generalizations can be considered applicable to the social and psychological world of proto-human higher primate species for a period of approximately 40 million years. Against this background, hominids evolved during the last few million years, culminating in the emergence of our species within the last few hundred thousand years, and finally in the appearance of truly modern Homo sapiens within the last hundred thousand. Aside from the increase in intelligence, as indicated by increasing relative and absolute brain size as well as by increasing complexity of stone tools, one hallmark of the transition to hominids was an increasing reliance on hunting. All monkeys and apes are largely vegetarians and insect-eaters; instances of meat-eating are instructive but relatively infrequent.
Among the most technologically primitive humans, whether in the fossil record or in the ethnographic one, hunting is invariably of major importance (Lee and DeVore 1968, Dahlberg 1981). Most of the stone tools that have survived archaeologically were sued in hunting or butchering, and the demands caused by this activity have long been held to be central to the emergence of human intelligence and social organization. Recent evidence has shown that the stone used for this purpose had to be traded over long distances, implying unexpectedly complex social networks among our ancestors before 2 million years ago. Furthermore,
even chimpanzees share meat after a kill (but not plant foods), and among human hunter-gatherers, the following of elaborate regulations for such sharing of meat may be a life-and-death matter. Finally, with one noteworthy exception (the Agta of the Philippines, where women routinely hunt), all hunting and gathering societies in the ethnographic record have a division of labor by sex, with men doing almost all the hunting and providing meat for their families and women providing most of the vegetable foods. These features of soceity related to hunting had to have been grafted onto an already complex social life characteristic of nonhuman higher primates.
However, this traditional male-centered view gives at most half the story (Dahlberg 1981). In many hunting and gathering societies, plant foods gathered by women constitute more than half of the diet. Plant foods are shared in these societies (although not beyond the immediate family), while they are not shared in nonhuman primates. The early advent of upright posture may have had more to do with the need for females to carry plant foods as well as infants to a base camp than with any advantage upright posture conferred in hunting, and it may be that among the first tools invented were digging sticks and carrying devices for plants or infants—tools that, however crucial, would not be preserved in archaeological assemblages and that would most likely have been invented by women.
Psychodynamic theorist John Bowlby coined the phrase "the environment of human evolutionary adaptedness," which aptly describes the hunting and gathering way of life. The phrase correctly implies that this is, or was, the context for which natural selection prepared us and from which we have departed only in the past 10,000 years, a very short time in evolutionary terms. (The industrial revolution, in the same terms, happened only a moment ago.) From many studies of recent hunting and gathering peoples, combined with archaeological evidence of those of the distant past, it is possible to make certain generalizations about this context (lee and DeVore 1968): social groups were usually small, ranging in size from fifteen to forty people related through blood or marriage; groups were nomadic, moving frequently to take advantage of changing subsistence opportunities, and flexible in composition, size, and adaptive strategies; daily life involved physical challenge, vigorous exercise, and occasional hunger but with a usually dependable food base from a moderate work effort and with a marked division of labor by gender; disease, mainly infectious rather than chronic, produced high rates of mortality especially in infancy and early childhood, with consequent frequent experience of loss; virtually all of life's activities were done in a highly social context with people one knew well, often the same people for different activities; and privacy was limited, but creative ex-
pression in the arts was frequently possible, and conflicts and problems were dealt with through extensive group discussions.
These generalizations describe the context in which most of human evolution and prehistory have occurred, so it is often said that we are, in effect, hunter-gatherers in skyscrapers. Simplistic observations about the consequences of this change are often made and are not helpful. Mental illnesses, both major and minor, are present in such societies. All of them experience some level of violent conflict, up to and including homicide. Other human behaviors usually considered undesirable, such as selfishness, deceit, adolescent rebellion, adultery, desertion, and child abuse, occur in such societies, and it is impossible to compare the rates of such behaviors to those in our own society. Although fluid in their way, life in such societies is restricted to a much smaller number of people than ours, and the lack of privacy and inability to get away from those people must constitute stresses, just as crowding and high levels of contact with strangers may constitute stress in advanced societies. The stresses of illness and mortality, as well as the stresses and uncertainties of the daily food quest, also must take their toll. A truly thoughtful set of observations and analyses of the differences between psychological conditions in our society and the kind of society in which we spent most of our history could be imagined but has not yet been made.
III
Since the late 1960s, there has been rapid emergence of an influential new field of evolutionary study known as neo-Darwinian theory or, more commonly, sociobiology (Wilson 1975, Trivers 1985). This new set of principles has been quickly adopted by most investigators who study animal behavior under natural conditions, including ethologists and behavioral ecologists, and has also influenced many anthropologists and psychologists. Briefly summarized, the principles are as follows (Trivers 1985):
An organism is, in essence, a gene's way of making another gene. Put more strictly, it is a way found by up to thousands of genes, though shortor long-term cooperation, to make copies of themselves. As long as it is admitted that there can be no forces operating in nature other than physicochemical ones, it must be admitted that continued membership in an ongoing germ plasm is the only goal served by any given gene. To the extent that a gene influences behavior, it can only continue in the germ plasm if it maintains or enhances, through the behavior, the number of copies of itself in the gene pool of the next generation. (Contrary to a frequently repeated confusion, the cohesiveness of the genome has only quantitiative, not qualitative, bearing on the validity of this Principle.)
Genes increase their number by enhancing reproductive success. Enhancing survival is only one way of doing this. Where the two goals, enhancing survival and enhancing reproductive success, are in conflict, as they often are, genes that enhance reproductive success will replace genes that enhance survival. The concept of fitness in evolutionary theory has no meaning except the relative frequency of characteristics and of the genes that influence them. It is a tautological description of reproductive success and has nothing necessarily in common with medical, social, or athletic definitions of fitness, all of which can be and often are achieved without an increase, or even with a decrease, in technically defined reproductive fitness.
Fitness is now best defined as inclusive . By inclusive fitness is meant the tendency of genes to influence their frequency not only through the survival and reproduction of the individual carrying them but also through the survival and reproduction of other closely related individuals who may be carrying the same genes through common descent. This is the concept introduced by Hamilton (1964) to account, using the mathematics of evolutionary genetics, for the existence of altrusim in animals, which previously seemed something to be culled by the process of natural selection. Hence the need for a newly defined subprocess of natural selection, called kin selection. If I die to save my identical twin, then the frequency of any gene that helped predispose me to that action will (all else being equal) be unaffected by my death. In general terms, such genes, or any genes predisposing me to self-sacrifice for a relative, should be favored under conditions where the ratio of the recipient's benefit to the altruist's costs exceeds the inverse of the degree of genetic relatedness. This concept has been invoked to explain self-sacrifice of soldier ants for the colony, alarm calls of birds and ground squirrels, and nepotism in human beings, among many other phenomena of human and animal behavior. Other theories brought to bear on the problem of altruism have been reciprocal altruism and the prisoner's dilemma model of cooperation, neither of which requires that the altruist and the recipient be related.
As argued by Robert Trivers (1972, 1985) from a suggestion of Darwin's, in species with two sexes in which there is a physiological or behavioral difference in the energy invested in individual offspring, the sex that invests more will become the scarce resource over which the other sex competes. In mammals and in most birds, females exhibit greater investment, but direct male parental investment may be very high in some species. Species in which male parental investment is high tend to be those in which pair-formation of a breeding male with the breeding female is long lasting; sexual dimorphism, both morphological and behavioral, is low; male competition for females is low; and variability among males in
reproductive success is low. These "pair-bonding" species, a category including 8,000 species of birds but only a minoirty of mammal species, may be contrasted with "lek" or "tournament" species, so called because they sometimes have annual seasonal breeding "tournaments" in which males compete fiercely for females. These species often have high sexual dimporphism for display or fighting (e.g., antlers or peacock feathers), low tendency for pair formation, low direct male parental investment in offspring, and high variability in reproductive success.
Human beings are considered to be near but not at the pair-bonding extreme of the continuum, as viewed from the perspective of sexual dimorphism, degress of direct male involvement in the care of offspring in a wide range of cultures, and the known distribution of human marriage forms (Daly and Wilson 1983, Murdock 1967). Polygyny, in which one man marries several women, is allowed or encouraged in 83 percent of known cultures in the anthropological record (708 of 849), while the converse arrangement, polyandry, is rare (4 of 849), and a double standard of sexual restriction is extremely common; still, most human marriages have probably been monogamous at least in intent.
A neo-Darwinian model of parent-offspring conflict advanced by Trivers (1974, 1985) has profound implications for the nature of the family. Weaning conflict is very common among mammals, and there are equivalent phenomena among birds, even including tantrum behavior on the part of the weanling. If the evolutionary purposes of mother and offspring were isomorphic, then they should "agree" (the use of this kind of language is shorthand for "should have been selected to act as if they agreed" and implies no conscious intent) that a given level and duration of investment is necessary and sufficient, after which attention should be turned by the mother to her next unborn offspring. A naive model of the nature of the family assumes that since it functions as a harmonious unit under ideal conditions, that is presumably how it was designed by evolution. However, it was not so designed. Like the breeding pair discussed above, it is an association among individuals with overlapping but distinct evolutionary purposes. Its members naturally pursue individual goals that are sometimes at odds with each other's ultimate (not merely temporary) purposes, and their relations are naturally conflictful rather than harmonious. This natural conflict is not the result of friction in what should be or could be a smoothly functioning system but is intrinsic.
Competition among unrelated individuals can be expected at times to be extreme. Virtually all animal species for which there is sufficent evidence have been seen to exhibit extremes of violent conflict, up to and including homicide, in the wild (Wilson 1975). The belief that human beings are rare among animal species in that we kill our own kind is erroneous, and more evidence to the contrary accumulates every year.
One particularly noteworthy phenomenon is competitive infanticide , already alluded to in relation to wild chimpanzees. The paradigmatic case is that of the Hanuman langur moneky of India, as observed by Sarah Blaffer Hrdy (1977), among others. Langur troops consist of a core of related females and their offspring, associated for a period of a few years or less with unrelated immigrant males. Occasionally, new males arrive and challenge the resident males. If they win and take over the troop, they drive the previous males away and proceed systematically to attack and kill resident infants under six months of age. The mothers of those infants then come into estrus again (much sooner than they would have if the infants has survived and continued to nurse) and are impregnated by the new males. Controversy has centered over whether this is normal behavor or a response to crowding or other soical stress. Such controversy misses the point that the behavior enhances the reproductive success of the new males at the expense of the old ones and therefore can be expected to be favored by natural selection, whether or not we choose to call it normal. Furthermore, similar phenomena have now been observed in many species.
The delineation of universal features of human behavior is central to the elucidation of the effects of phylogeny. But natural selection operating on organisms ancestral to ourselves created not only individuals with certain needs, drives, and capacities but also equations—"if-then" statements—relating the environment to the social system, the social system to the individual, and so on. In a cross-cultural study of husband-wife intimacy (Whiting and Whiting 1975), it was hsown that societies in which husbands eat, sleep, and care for children together with their wives are also those that are less belligerent. That is, phylogeny appears to have provided a system such that separating men from women and small children is associated with successful adapation as warriors. This is not to say that they must be warriors or that they must be aloof from their wives but that choosing aloofness may increase effective belligerency and/orperhaps vice versa. The universal characteristic here is not a phenotypic characteristic at all but an underlying mechanism relating two sets of characteristics to each other. (An extended discussion of this conceptual framework may be found in Konner 1982b .)
In the past decade, the application of neo-Darwinian or sociobiological theory to ethnological materials has produced many findings that seem to bypass the complex questions of the relationships among society, cutlure, and individual development. For example, societies in which young men inherit land from their mothers' brothers are more lax about the prevention of female adultery than are societies in which young men inherit from their fathers (Kurland 1979); in societies in which polygyny
is allowed, wealthier and more influential men tend to have more wives (Betzig 1986); and in small-scale societies in which adoption of children it common, it tends to follow patterns predicted by genetic relatedness (Silk 1987). Investigators mkaing these findings usually declare that they do not claim any direct genetic basis for these variations in human behavior, and indeed some of the worst confusion about sociobiology stems from a failure to appreciate this distinction between the propositions of neo-Darwinian theory and those of traditioanl behavioral genetics or molecular genetics.
Even in a nonhuman species such as the red-winged blackbird, males singing on richer territories mate with several females instead of one (Harding and Follett 1979). But the mechanism of this flexible adaptive system—known as a facultative adaptation—must be quite different in blackbirds than the similar mechanism in human beings (although it would probably underestimate blackbirds to assume that in them the system is under tigth genetic control). The wings of insects some from thoracic tissue, the wings of birds from forearm structures, the wings of bats from fingers, and the wings of humans from technology. These four adaptations to the problem of flight serve similar functions with extremely different provenance. The same will prove to be true of adaptations in social behavior (Konner 1982b ).
Neo-Darwinian or sociobiological theory is sometimes presumed to carry value judgments as a necessary concomitant. This logic appears to suggest that whatever is, is right. An extension of this fallacy would hold that sickle cell anemia and thallasemia must be accepted because natural selection has maintained them through balnced polymorphism, the heterozygotes being at an advantage in malaria resistance; or that myopia should not be corrected because natural selection in favor of sharp vision has relaxed in the human population since the end of the hunting and gathering era. Human judgments about what is desirable are separate from and must take precedence over any observations or explanations of what exists in nature, although they may be enhanced by taking the facts of the natural world into account.
IV
The main enterprise of cultural anthropology has been the description and analysis of cross-cultural variation, but the enterprise has always had an inevitable, even if tacit, complement: the characterization of features of human behavior that do not vary or that vary relatively little. The concept of universals has, howeve,r at least five different meanings: behaviors, such as coordinated bipedal walking or smiling in social greeting, that are exhibited by all normal adult members of every known
society; behaviors that are universal within an age or sex class, such as the refleces in all normal neonates or the ejaculatory motor action pattern in all postpubertal males; population characteristics that apply to all populations but not all individuals, such as the sex difference in physical aggressiveness; universal features of culture rather than of behavior, such as the taboos against incest and homicide, or the highly variable but always present institution of marriage, or the social construction of illness in attempts at healing; and, finally, characteristics that, although unusual or even rare, are found at some low level in every population, such as homicidal violence, thought disorder, depression, suicide, or incest.
The list of characters that fill these five categories is a very long one, much longer than the prominent anthropologists of the early heyday of cross-cultural study would have predicted (Eibl-Eibesfeldt 1988). The search for societies without violence, or without gender differences that go beyond childbearing, or without mental illness, or even without the ability to make and use fire has been a vain one, and although there is convincing documentation of variation in the incidence or context of expression of most human behaviors, the existence of such a large core of constantly present, if variable, features constitutes a demonstration of the reality of human nature and its validity as a scientific construct (Konner 1982b ).
In the realm of psychosocial development, Freud postulated, and present-day child psychiatry continues to accept in altered and disputed forms, a universal sequence of emotional development on which the social environment of the family could be claimed to produce enduring traits of emotional disposition. Beyond some very general elements—the existence of infantile sexuality, the formation of an attachment to a primary caretake who is usually the mother, the ubiquity of conflicts and jealousies within the family—this allegedly universal sequence has never found empirical support; hence the unresolvable disputes among different schools of psychoanalysis and the enduring skepticism of outsiders. Extensive cross-cultural studies of human behavioral and psychological development have not produced evidence relevant to these particular models, but they have produced extensive evidence supporting some more empirically grounded putative universals of psychosocial growth. In the absence of knowledge of neuropsychological development, psychoanalytic theory postulated a libidinal theory of this development (Freud 1920) that few take seriously today. However, the growing body of actual knowledge about neural and neuroendocrine deveopment can now begin to serve a parallel function in relation to the more empirically grounded newer studies of psychosocial growth.
Among the well-established cross-cultural universals of psychosocial development, the following are the best supported and in many cases can
be plausibly related to putative underlying neural events (Konner 1982a , 1991): the emergence of sociality, as heralded by social smiling, during the first four months of life, in parallel with the maturation of basal ganglia and cortical motor circuits; the emergence of strong attachments, as well as of fears of separation and of strangers, in the second half of the first year of life, in parallel with the maturation of the major fiber tracts of the limbic system; the emergence of language during the second year and after, in parallel with the maturation of the thalamic projection to the auditory cortex among other circuits; the emergence of a sex difference in physical aggressiveness in early and middle childhood, with males on the average exceeding females, a consequence in part of prenatal and rogenization of the hypothalamus; and the emergence of adult sexual motivation and functioning in adolescence, in parallel with and following the maturation of the reproductive system at puberty, against the background of the previously mentioned prenatal androgenization of the hypothalamus. In addition, there are a number of apparent cross-cultural universals of cognitive development.
There are other probable cross-cultural developmental universals, such as increased babbing in the second half of the first year and possibly progress through the first three, or perhaps four, of the six stages in Lawrence Kohlberg's scheme of moral development in childhood. Although the underlying neuropsychology is at an early stage of elucidation, the cross-cultural universality of these traits is well established; in each case, there is extensive experimental evidence to support the maturational nature of the process in behavioral development. They constitute a first approximation of the true structural basis of psychosocial development, which Freud groped fro whit his crude theory of libidinal development in the nervous system.
They also constitute a firm basis for the future understanding of how variations in social experience, whether clinical or cross-cultural, act on the maturing psychosocial competence to produce potentially lasting variations. In each of the processes mentioned, cross-cultural differentiation of the maturing competence begins almost as soon as the maturation occurs. However, the acceptance and increasingly detailed and reliable description of the maturational constants underlying the variation in psychosocial growth will provide a steadily firmer place on which to stand while attempting to investigate the role of cultural and individual experience.
V
Within the last two or three years, there has emerged a new mode of inquiry in the study of human nature. The current revolution in mo-
lecular genetics is rapidly leading both to a molecular neurogenetics and to a molecular genetics of behavior and mind. Its established empirical base already extends from the simplest animal systems to the most complex functions of the human mind and from the earliest to the latest stages of the life cycle. Some highlights among its recent discoveries are the following:
1) a family of genes coded for a set of neuropeptides that, in turn, controls the stereotyped egg-laying behavior of the sea slug Aplysia californica (Scheller et al. 1984);
2) a Drosophila behavioral mutant has been explained as the result of known genes that alter the potassium channel of the neuronal membrane (Tanouye et al. 1986);
3) a single-gene form of human pseudohermaphroditism has been traced to deficiency in 5-alpha-reductase, which converts testosterone to dihydrotestosterone; affected individuals have an ambiguous or feminine body habitus and psychological disposition until puberty, when they become physically and psychologically masculine despite having been raised as girls (Imperato-McGinley et al. 1979, 1980);
4) Huntington's disease, a neuropsychiatric condition that presents in the thirties or forties (often beginning with psychiatric manifestations), has been shown to be caused by a dominant gene on the short arm of chromosome 4 (Gusella et al. 1984);
5) manic-depressive illness (bipolar affective disorder), a severe form of mental illness presenting in adulthood, has been shown in different kindreds to be caused in part by a gene on the X chromosome (Baron et al. 1987);
6) Alzheimer's disease, the most common form of late-life dementia, has been shown to be caused in some kindreds by a gene on chromosome 21; a DNA probe cloned from the amyloid protein long known to accumulate in the brains of affected people has been localized to the same chromosome section; and the same gene is duplicated in individuals with Down's syndrome—who develop early Alzheimer's—even in the absence of the usual chromosomal anomaly, a duplication of the whole chromosome (Tanzi et al. 1987; St. George-Hyslop et al. 1987; Goldgaber et al. 1987; Delabar et al. 1987).
These recent findigns remove any doubt regarding the ability of genes to alter complex behavior through comprehensible biochemical pathways affecting the nervous system. They are not subject to any of the methodological criticisms always leveled at classical Mendelian behavioral genetics, especially at correlational human studies. Extreme skeptics will still claim that these findings have no bearing on the development of normal human behavior, but there is no reason to believe that they are right. On the contrary, it is likely that of the hundred thousand
or so genes in the human genome, the ones most easily identified would be the ones with the most devastating effects and that subsequently it would become possible to identify genes affecting physiology and behavior within the normal range. We can expect such identifications within one or two decades.
Such findings, however, are not the end but the beginning of the development of the epigenetic—interactionist—approach to human nature. In the absence of other treatments, few would argue that we should refrain from "gene surgery" for Alzheimer's disease, but even with manic-depressive illness there will be serious ethical issues in tampering with the system, which may produce certain desirable personality variations and even exceptional creativity as well as serious mental illness. Within the normal range, the arguments for restraint would be even stronger. Nevertheless, from the viewpoint of analysis of the development of behavior, a technology of molecular genetics will provide insights that will transform our view of human nature. Already we have learned that both cognitive and emotional functions can be altered by specific genes and that genes can manifest themselves in the behavioral phenotype at almost any stage of life, sometimes without prior developmental manifestations of any kind. The question is, How will the environment prove to interact with genetic predispositions and vulnerabilities?
Phenylketonuria provides a valuable model, It is a simle genetic disorder with increasingly understood molecular genetics, yet with an environmental proximate cause and a solution hat involves modification of one of the most mundane features of the environment, the diet. Such environmental modification essentially obviates the impact of the genotype. Drug treatments have also bee tried, and in the not too distant future, a molecular genetic intervention in the germ line ("gene surgery") may be possible. At that point, there will be a full spectrum of treatment possibilities, making phenylketonuria a paradigmatic instance of the complex interactions of genotype, metabolism, and environment in the development of neural, behavioral, and mental phenomena. It is not difficult to imagine in broad outline the similar range of interpretations that will develop for a variety of other abnormalities of brain and behavior and indeed for some variations within the normal range.
In addition, there are likely to be many new discoveries about the way environment exerts its influence on the development of behavior, and some of these are likely to be surprising. Neuroembryological research has given new meaning to the concept of plasticity by showing the great importance of competition among neurons and connections for destination sites in the developing brain, and the process of selective pruning goes on postnatally as well as prenatally (Changeux 1985; GoldmanRakic et al. 1983). Brain functions as crucial to psychology as laterality
may depend on complex intrauterine mechanisms under indirect, rather than direct, genetic control (Geschwind and Galaburda 1987), and these mechanisms of plasticity were not only unexplored but unsuspected until a few years ago.
VI
In the eithteenth century, Goethe (Magnus 1949) posited the existence of a fundamental or archetypal form in plant life, a sort of key to the order of life itself in the largest sense, and perhaps even of the universe beyond the realm of the living. Indeed, he made his search for order a foil to that of Newton, which he considered too mechanical; he thought that even the fundamental ordering principles of the universe might be biological, not physical. Although in this he went too far, there is a sense in which the Urpflanze , Goethe's ultimate plant form, really does exist and in which it has become, two centuries after he posited it, as central to the enterprise of at least the life sciences as he believed it would be.
The Urpflanze is, of course, DNA, and with the scientific unraveling of its form has come, and will continue to come, a sense of order and power in the realm of biology that only a mind like Goethe's could have imagined. The extent to which the realm of behavior will also come under the sway of this intellectual order remains to be seen; but even the answer to this question will be largely provided by that order. Undoubtedly, the answer will involve mechanisms of epigenesis that include such phenomena as operant conditioning, snesitive periods, psychological trauma, cognitive maps, and symbol systems as well as diet, infection, and injury. But the way these phenomena operate—within the constraints provided by the human genome—to produce individual and collective behavioral patterns and tendencies is vastly more uncertain than the textbooks in the relevant fields allow, and the mutual contradictions of those textbooks underscore the uncertainties. That the delineation of how the environment affects the individual, beyond the hoary pieties of plasticity, remains a task almost completely for the future is perhaps the greatest discovery of the past twenty years.
Two further implications of recent advances in biology need to be emphasized. First, if neo-Darwinian principles of behavior and reproduction are even mostly correct, the fundamental metaphor of modern social science is in error. That metaphor, which is as venerable as social science itself, claims that society is an organism, with individuals as cells, specialized subgroups of individuals as tissues or organs, and conflict as a transient aberration, or pahtology, the elimination of which restores the social organsm to health. The basic weakness of the analogy is most starkly exposed each time an individual or group departs from the soci-
ety and joins or forms another—something the cells or organs of an animal cannot do, of course—but in fact the weakness is evident in the ubiquity of social conflict even within the most intimately interdependent social relationships. Such conflict is not inadvertent friction in a system that should, by design, function smoothly but is an inherent and inevitable expression of the purposes of social life itself.
Second, the motivational portions of the brain, particularly the hypothalamus, have functional characteristics relevant to the apparent chronicity of human dissatisfaction. Animal experiments on the lateral hypothalamus suggest that the motivated condition is to some extent nonspecific, with the internal state responsive to but not geared for the particular external circumstances. A continuum between attentiveness or alertness and intense drive states ensures that responsiveness will never be long delayed but also that it will not always be appropriate and, more important, that the organisms's chronic internal state will be a vague mixture of anxiety and desire—best described perhaps by the phrase "I want," spoken iwth or without an object for the verb. This insight of physiological psychology about the internal motivational states of animals like ourselves fits well with the more recent insights of sociobiology about the conflictful external relations entered upon by the same and similar animals and also with the failure of the organismal model of societal coherence.
One consequence of these insights is that the view of life current in behavioral biology bears more resemblance to the view taken in the time-honored traditions of the humanities than either does to the canons of social science. Henry James once described life as "slow advance into enemy territory" and wrote, "Life is , in fact, a battle, Evil is insolent and strong; beauty enchanting, but rare; goodness very apt to be weak; folly very apt to be defiant; wickedness to carry the day; imbeciles to be in great places, people of sense in small, and mankind generally unhappy" (Zeibel 1951). Similar sentiments have been common in the literary traditions of many societies from the earliest religious and epic sagas to novels and plays completed this morning. Religious traditions of varied character recognize the reality of a deeply, even tragically, flawed human nature, but they exhort against it, while literary artists seem satisfied to describe it. In either case, it is viewed, sadly, as all too real, and these vividly brilliant clasic observations of it fit well with Darwin's remark to Joseph Hooker: "What a book, a Devil's Chaplain might write on the clumsy, wasteful, blundering low and horribly cruel works of nature!"
Of course, that nature also includes an equally inherent ethical component that derives from the necessity for cooperation and altruism, the potential for responsibility, decency, love, even happiness. These capacities, too, are shared by amny other animals, and we can take encouragement from the fact that they are so widespread in nature. But for them
to prevail requires the kind of collective attention that is possible only in the framework of human culture. In this framework, reflection on the outcomes of natural tendencies results in judgments that restrain or modify those tendencies. It is full of deceptions, but it is much better than nothing, and it exceeds the capabilities of any other animal for similar restraint and modification.
The evolutionary, biological view of human nature provides many parallels with that of animal natures and only a few clear distinctions. Traditionally and presently, distinctions between ourselves and animals have emphasized the primacy and complexity of human rational faculties. But in recent years, the development of artificial intelligence has duplicated a surprising number of those same faculties, and in the community of people who think about the implications of this fact, it is common to distinguish humans from hacines by referring to the human emotional faculties—precisely those we share in common with animals. It would seem that we are sorted to a pulp, caught in a vise made, on the one side, fo the increasing power of evolutionary biology in explaining the emotions and, on the other, of the relentless duplication of human mental faculties by increasingly suble and complex machines. So, what is left of us?
What is left is that only we combine the emotions and the life cycle drama of the animal world with a fully empowered reflective and communicative faculty. No other animal has that faculty, and no machine has an animal bodily life. Other animals can communicate, but they do not exchange views on the rightness or wrongness of their emotions. Machines can network and think, but they cannot discuss their fear of dying. What religious people think of as the soul or spirit can perhaps be fairly said to consist of just this: the intelligence of an advanced machine in the mortal brain and body of an animal. And what we call culture is a collective way of using that intelligence to express and modify the emotions of that brain, the impulse and pain and exhibaration of that body. Both the intelligence and the impulse, the communicative capability and the pain, are components of human nature, and the wy they interact is the unique feature of that nature. Without conceding the existence of human nature, without describing it as forthrightly and richly as possible, we will never fully exercise that crucial feature, which alone holds the prospect of an admittedly limited but absolutely imperative transcendence.
References
Arling, G. L., and H. F. Harlow, "Effects of Social Deprivation on Maternal Behavior of Rhesus Monkeys," Journal of Comparative and Physiological Psychology 64 (1967): 371-377.
Baron, M., N. Risch, R. Hamburger, B. Mandel, S. Kushner, M. Newman, D. Drumer, and R. H. Belmaker, "Genetic Linkage between X-chromosome Markers and Bipolar Affective Illness," Nature 326 (1987): 289-272.
Barry, H., III, and L. Paxson. "Infancy and Early Childhood: Cross-cultural Codes," 2. Ethnology 10 (1971).
Betzig, L. L. Despotism and Differential Reproduction: A Darwinian View of History. New York: Aldine, 1986.
Boslby, J. Attachment and Loss. 3 vols. London: Hogarth Press, 1969-1977.
Changeux, J.-P. Neuronal Man: The Biology of Mind. New York: Oxford University Press, 1985.
Cheney, D., R. Seyfarth, and B. Smuts. "Social Relationships and Social Cognition in Nonhuman Primates," Science 234 (1986): 1361.
Ciochon, R. L., and J. G. Fleagle, eds. Primate Evolution and Human Origins. New York: Aldine de Gruyter, 1987.
Dahlberg, Frances, ed. Woman the Gatherer. New Haven: Yale University Press, 1981.
Daly, Martin, and Margo Wilson. Sex, Evolution, and Behavior. 2d ed. Boston: Willard Grant, 1983.
Delabar, M., D. Goldgaber, Y. Lamour, A. Nicole, J. Huret, J. De Grouchy, P. Brown, D. C. Gajdusek, and P. Sinet. "b Amyloid Gene Duplication in Alzheimer's Disease and Karyotypically Normal Down Syndrome," Science 235 (1987): 1390-1392.
Eibl-Eibesfeldt, I. Human Ethology. New York: Aldine Press, 1988. (Translation of Die Biologie des Menschlichen Verhaltens: Grundriss der Humanethologie. Munich: Piper, 1984.)
Freud, S. A General Introduction to Psychoanalysis. New York: Washington Square Press, 1920.
Galaburda, A. M., and T. L. Kemper. "Cytoarchitectonic Abnormalities in Developmental Dyslexis: A Case Study," Ann. Neurol. 6 (1979): 94-100.
Geschwind, N., and A. M. Galaburda. "Cerebral Lateralization: Biological Mechanisms, Associations, and Pathology: A Hypothesis and a Program for Research, Pts. I, II, III. Arch. Neurol. 42 (1985): 428-459, 521-552, 634-654.
———. Cerebral Lateralization. Cambridge: MIT Press, 1987.
Goldgaber, D., M. I. Lerman, O. W. McBride, U. Saffiotti, and D. C. Gajdusek. "Characterization and Chromosomal Localization of a cDNA Encoding Brain Amyloid of Alzheimer's Disease," Science 235 (1987): 877-880.
Goldman-Rakic, P. S., A. Isseroff, M. L. Schwartz, and N. M. Bugbee. "The Neurobiology of Cognitive Development," in P. H. Mussen, M. M. Haith, and J. J. Campos, eds., The Handbook of Child Psychology. Vol. 2: Infancy and Developmental Psychobiology. New York: John wiley & Sons, 1983, 281-344.
Goodall, J. The Chimpanzees of Gombe: Patterns of Behavior. Cambridge: Harvard University Press, 1986.
Gusella, J. F., R. E. Tanzi, M. A. Anderson, W. Hobbs, K. Gibbons, R. Raschtchian, T. C. Gilliam, M. R. Wallace, N. S. Wexler, and P. M. Conneally. "DNA Markers for Nervous System Diseases," Science 225 (1984): 1320-1326.
Hamilton, W. d. "The Genetical evolution of Social Behavior, I & II," Journal of Theoretical Biology 7 (1964): 1-16, 17-52.
Harding, C. F., and B. K. Follett. "Hormone Changes Triggered by Aggression in a Natural Popultion of Blackbrids," Science 203 (1979): 918-920.
Harlow, H. F. "The Maternal Affectional System," in B. M. Foss, ed., Determinants of Infant Behavior. Vol. 2. London: Methuen, 1963.
———. "Age-mate or Peer Affectional System," in D. Lehrman, R. Hinde, and E. shaw, eds., Advances in the Study of Behavior. Vol. 2. New York: Academic Press, 1969.
Hrdy, S. B. The Langurs of Abu: Female and Male Strategies of Reproduction. Cambridge: harvard University Press, 1977.
Imperato-McGinley, J., R. E. Peterson, T. Gautier, and E. Sturla. "Androgens and the Evolution of Male-Gender Identity among Male Pseudohermaphrodites with 5a-Reductase Deficiency," The New England Journal of Medicine 300 (1979): 1233-1270. Also in S. Chase and A. Thomas, eds., Annual Progress in Child Psychiatry and Child Development. New York: Brunner/Mazel, 1980.
Kagan, J. "The Uses of Cross-Cultural Research in Early Development," in T. H. Leiderman, S. R. Tulkin, and A. Rosenfeld, eds., Culture and Infancy. New York: Academic Press, 1977.
—–—. The Nature of the Child. New York: Basic Books, 1984.
Kagan, J., and R. E. Klein. "Cross-Cultural Perspectives on Early Development." American Psychologist 28 (1973): 947-961.
Konner, M. J. "Evolution of Human Behavior Development," in R. L. Munro, R. Munnro, and B. Whiting, eds., Handbook of Cross-Cultural Development. New York: Garland Press, 1981.
———. "biological Aspects of Mother-Infant Bond," in R. Emde and R. Harmon, eds., Development of Attachment and Affiliative Processes. New York: Plenum, 1982a .
———. The Tangled Wing: Biological Constraints on the Human Spirit. New York: Holt, Rinehart, Winston, 1982b . (Harper & Row paperback, 1983.)
———. "Universals of Psychosocial Growth in Relations to Brain Myelination," in A. Petersen and K. Gibson, eds., Brain Development and Social Behavior. New York: Aldine-DeGruyter, 1991.
Kurland, J. A. "Paternity, Mother's Brother, and Human Sociality," in N. A. Chagnon and W. Irons, eds., Evolutionaary Biology and Human Social Behavior. North Scituate, Mass.: Duxbury, 1979, 145-180.
LeBoeuf, B., and R. S. Peterson. "Social Status and Mating Activity in Elephant Seals," Science 163 (1969): 91-93.
Lee, R., and I. DeVore. Man the Hunter. Chicago: Aldine, 1968.
Maccoby, E. E., and C. N. Jacklin. The Psychology of Sex Differences. Stanford: Stanford University Press, 1974.
MacLean, P. D. "Brain Evolution Relating to Family, Play, and the Separation Call," Arch. Gen. Psychiatry 42 (1985): 405.
Magnus, r. Goethe as a Scientist. New York: Henry Schuman, 1949.
Mason, A. J., J. S. Hayflick, R. T. Zoeller, W. S. Young, III, H. S. Phillips, K. Nikolics, and P. H. Seeburg. "A Deletion Truncating the Gonadotropin-Releasing Hormone Gene Is Responsible for Hypogonadism in the hpg Mouse," Science 234 (1986): 1366-1371.
Mason, A. J., S. L. Pitts, K. Nikolics, E. Szonyi, J. N. Wilcox, P. H. Seeburg, and T. A. Stewart. "The Hypogonadal Mouse: Reproductive Functions Restored by Gene Therapy," science 234 (1986): 1372-1378.
McKusick, V. Mendelian Inheritance in Man. 7th ed. Baltimore: Johns Hopkins University Press, 1986.
Munroe, R. H., R. L. Munroe, and B. B. Whiting. Handbook of Cross-Cultural Human Development. Chicago: Garland STPM Press, 1981.
Murdock, G. P. Ethnographic Atlas: A Summary. Pittsburgh: University of Pittsburgh Press, 1967.
Napier, J., and P. H. Napier. The Natural History of the Primates. Gambridge: MIT Press, 1985.
Nemiah, J. "chapter 20: The Neuroses," in H. Kaplan and B. Sadock, eds., Comprehensive Textbook of Psychiatry. 4th ed. Baltimore: Williams and Wilkins, 1985.
Plomin, R., J. C. DeFries, and G. E. McClearn. Behavioral Genetics: A Primer. San Francisco: Freeman, 1980.
———. Development, Genetics, and Psychology. Hillsdale, N.J.: Lawrence Erlbaum Associates, 1986.
Sackett, G. P., G. c. Ruppenthal, C. E. Fahrenbruch, and R. A. Holm. "Social Isolation Rearing Effects in Monkeys Vary with Genotype," Developmental Psychology 17 (1981): 313-318.
Scarr, S., and K. K. Kidd. "Developmental behavioral Genetics," in P. H. Mussen. M. M. Haith, and J. J. Campos, eds., The Handbook of Child Psychology. Vol. 2: Infancy and Developmental Psychobiology. New York: John Wiley & Sons. 1983, 345-433.
St. George-Hyslop, P. H., R. E. Tanzi, R. J. Polinsky, J. L. Haines, L. Nee, P. C. Wtkins, R. H. Myers, R. G. Feldman, D. Pollen, D Drachman, J. Growdon, A. Bruni, J. Foncin, D. Salmon, P. Frommelt, L. Amaducci, S. Sorbi, S. Placentini, G. D. Stewart, W. J. Hobbs, P. M. Conneally, and J. F. Gusella. "The Genetic Defect Causing Familial Alzheimer's Disease Maps on Chromosome 21," Science 235 (1987): 885-890.
Scheller, R. H., R. Kaldany, T. Kreiner, A. C. Mahon, J. R. Nambu, M. Schaefer, and R. Taussig. "Neuropeptides: Mediators of Behavior in Aplysia," Science 225 (1984): 1300-1308.
Shostak, M. Nisa: The Life and Words of a !Kung Woman. Cambridge: Harvard University Press, 1981.
Silk, J. "Adoption and Fosterage in Human Societies: Adaptations or Enigmas?" Cultural Anthropology 2 (1987): 39-49.
Smuts, B. Sex and Friendship in Baboons. New York: Aldine, 1985.
Stoller, R. J., and G. H. Herdt. "theories of Origins of Male Homosexuality," Arch. Gen. Psychiatry 42 (1985): 399.
Tanouye, M. A., c. A. Kamb, L. E. Iverson, and L. Salkoff. "genetics and Molecular Biology of Ionic Channels in Drosophila," Annual Review Neuroscience 9 (1986): 255-276.
Tanzi, R. E., J. F. Gusella, P. C. Watkins, G. A. P. Bruns, P. St. George-Hyslop, M. L. Van Keuren, d. Patterson, S. Pagan, D. M. Kurnit, and R. L. Neve.
"Amyloid b Protein Gene: cDNA, mRNA Disribution, and Genetic Linkage near the Alzheimer Locus," Science 235 (1987): 880-884.
Trivers, R. L. "Parental Investment and Sexual Selection," in B. Campbell, ed., Sexual Selection and the Descent of Man, 1871-1971. Chicago: Aldine, 1972.
———. "Parent-Offspring Conflict," american Zoologist 14 (1974): 249-264.
———. Social Evolution. Menlo Park: Benjamin Cummings, 1985.
whiting, J. W. M., and B. B. Whiting. "Aloofness and Intimacy between Husbands and Wives," Ethos 3 (1975): 183.
Wilson, E. O. Sociobiology: The New Synthesis. Cambridge: Harvard University Press, 1975.
———. On Human Nature. Cambridge: Harvard University Press, 1978.
Zeibel, Morton, ed. The Portable Henry James. New York: Viking, 1951, 27.
Six—
Reflections on Biology and Culture
John Dupré
I shall concentrate here on Melvin Konner's essay (chap. 5) for a reason perhaps embarrassingly characteristic of professional philosophers: I disagree with a great deal of it. However, Arnold Davidson, Harriet Ritvo, and Evelyn Fox Keller all help to illustrate central points underlying my dissent from some of Konner's claims, so I shall briefly try to bring out some of these general points before turning to my main task of dissent.
Let me begin with some very general remarks that apply to both Ritvo's and Davidson's contributions. As a general theoretical proposition, it would now be quite widely agreed that how we understand both ourselves and nature must depend considerably on our general theoretical beliefs. Despite major differences in subject matter and approach, I take both Ritvo and Davidson to be offering, among other things, more specific illustrations of this general truth.
But it is not just the understanding of, or, in the case of Davidson's subject matter, the affective response to, phenomena that is liable to such influences but even what we take to be the phenomena themselves. Many of Davidson's monsters, I take it, were quite widely supposed to actually exist, and their existence offered a variety of explanatory etiologies. The unpleasant demise of Davidson's masturbators is not merely asserted but described in amazing detail. In Ritvo's sources, the importance of bovine virtue is not merely assumed, detailed "factual" reports of the alarming consequences of its absence are offered. The "telegonic" effects of previous sexual partners on female animals are illustrated with numerous anecdotal instances.
These observations provide a useful background for my main focus: How far do such historical observations mandate skepticism about our
current theories and even alleged facts? In particular, how much skepticism is properly suggested about our current theories about animals and the consequences of these for our theories of ourselves as animals? The answer, I shall suggest later, is a good deal.
Ritvo's essay not only suggests the appropriateness of such skepticism but also indicates a particular ground. The tendency to see views of ourselves reflected in nature, the "projections" she so clearly documents, has also been suggested for more contemporary theories—as, indeed, we can see in Keller's essay, to which I shall return shortly.
There are obvious difficulties in relating historical lessons to critiques of contemporary beliefs. The opinions that Ritvo and Davidson recount strike us as ludicrous and either comical or grotesque. We have our own theories that we take to be if not amply demonstrated, as least sufficiently so to reveal the complete untenability of these historical views. Consequently, the conclusion that these views are projections of prejudices about ourselves onto animals has no serious contender. When we come to consider contemporary beliefs, however, particularly those that carry our highest epistemological honorific, "scientific," we have no such alternative and accepted standpoint from which to launch such a critique.
But as Keller has persuasively illustrated, this does not mean that clear grounds for such a critique cannot, nevertheless, be provided. I think there is considerable force to the objections Keller raises to contemporary evolutionary theory. Since I cannot possibly recapitulate the intricacies of her argument here, I shall make just one further observation in support of her conclusion. Contemporary theorists of the evolution of social behavior often assert that the central problem in their field is, or if they believe it to have been solved, was, the possibility of altruism. (Needless to say, altruism is a concept like competition that allows considerable equivocal play between technical and colloquial usages.) There are, of course, compelling reasons internal to evolutionary theory for perceiving altruism as a major problem. Nevertheless, if one is at all inclined to skepticism about the absolute objectivity of scientific theory, this must surely add plausibility to the contention that evolutionary theory recapitulates a traditional image of Hobbesian man.
There remain major questions that, I think, Keller's account raises but does not answer. In particular, I have in mind questions about specifically, I see two rather different, though perhaps not incompatible, sources of this individualism. Keller's main suggestion, in line with a point emphasized earlier in these remarks, is that we project views of human society onto the animal world; views, that is, of Hobbesian man or now, perhaps, Homo economicus var. Chicagoensis. However, the individualism she describes is also an example of what I take to be a more general method-
ological prejudice in science. I have in mind reductionism, or the general belief that the behavior of complex entities must be understood whooly in terms of the properties of their constituent parts.[1] Perhaps this methodological assumption should itself be understood as a more far-reaching projection of social philosophy onto the entirely of nature. If not, the relation between these positions is something that will require further exploration for a fuller understanding of these issues.
Keller, to return to my main thread, provides strong evidence that the dangers of projection of social ideology onto the natural world have relevance to our evaluation of contemporary as well as historical ideas. Konner, by Contrast, has a very different kind of project embodying a more complacent view of science: he encourages us to understand ourselves better by applying to ourselves the lessons that we have learned from our study of other animals. Since I take it we may all agree that we are, among other things, animals, this suggestion surely has something to commend it. However, the work of Ritvo and Keller should make clear a major danger inherent in such a project. To whatever extent our understanding of animals does involve a projection onto them of antecedent views about ourselves, the attempt to understand ourselves by applying our knowledge of naimals will complete a small and unilluminating circle. (I think there is a close analogy here with Terry Winograd's cautions about the use of machines as a route to understanding ourselves.) In fact, I believe that it is just this methodological problem, rather than the often very confused debate about the existence of "human nature," that is central to the accusation that human sociobiology, the most conspicuous contemporary attempt to illuminate human nature by appeal to more general biological theory, functions, in large part, as a rhetorical device for the defense of conservative ideology.[2]
As I have already confessed, I disagree with a good deal of Konner's argument. Even if this reflects no more than my own ideological biases, that itself would provide some evidence that even the evaluation or up-to-date scientific ideas is susceptible to such bias. Konner presents his views with subtlety and sophistication, and I certainly cannot hope, in the space of these comments, to give a comparable defense of an opposing position. What I shall do, however, is briefly indicate three major points at which I disagree strongly with the views he expresses.
First, Konner asserts at the outset that "an organism is in essence a gene's way of making another gene." This is, of course, an extreme instance of the reductionism I mentioned above in discussing Keller's contribution.[3] I also think it is profoundly misleading, even false. The issue here are complex and somewhat technical, and I can do the least justice to this part of my dissent. I take the fundamental error to be the assumption that a gene, or even, for that matter, an organism (as keller has argued),
can be adequately characterized in isolation. Or more accurately, the gene that is characterized in temrms of molecular biology cannot also be adequately described in terms of its phenotypic effects. Put simply, this is because the development of an organism involves enormously complex interactions between genes and with the environment, such that the question, what is the developmental upshot of a particular piece of DNA (such as an aquiline nose, or criminal tendencies)? is thoroughly ill-conceived.[4]
Konner is, of course, aware of these difficulties and remarks parenthetically that it is a confusion to suppose that the cohesiveness of the genome—that is, the enormously complex set of interactions between different genes and the environment in the development of an organism—has any but a quantitative bearing on the validity of this reductionistic premise. I believe he is mistaken. One way of seeing way this is so is to observe that given the incommensurability of the chemical and functional descriptions of genes, it is unclear that there is any interpretation of the term gene which could serve for an elaboration of Konner's Butlerian statement of the essence of an organism. As a matter of fact, this reductionistic slogan is generally offered as a consequence of speculations about the evolutionary process. But I think it is clear that if it cannot even be given ontogenetic sense, it has no chance at all of having a correct phylogenetic interpretation.[5] And finally, even if it were correct from both a developmental and en evolutionary perspective, there is a great deal more to an organism, especially a human one, than its ontogeny and phylogeny. I shall take up this remark again briefly at my conclusion.
My second main point ultimately concerns the role of language in science stressed by Keller and brings us back once more to the circle of projections with which I began this part of my commentary. Konner offers as support for the importance of biological considerations in understanding human behavior the claim that cultural anthropology has already discovered numerous cross-cultural universals. I suggest, on the contrary, that these divide exhaustively between the highly dubious and the irrelevant. In the latter category, I include examples such as coordinated bipedal walking and the reflexes of neonates. Certainly, I do not want to deny that human behavior is built on a substrate of (moderately) universal biological capacities. What are cleary more interesting are the universalistic claims about complex behaviors, and such claims I take to be uniformly dubious.
Let me illustrate the central problem with just one of Konner's examples. While admitting that the form is variable, he suggests that one significant universal—and hence presumably one that is to be taken to be grounded in biology—is the existence of marriage. But the variability of form, surely, is just the point. The question that needs to be asked is,
What is meant here by the word marriage ? Certainly for us, in modern Western society, the term is weighed down with social, economic, and affective connotations. Without a clear statement of which of these are, and which are not, included in the alleged anthropological universal, such a claim can do nothing but mislead. Any sense of the world marrriage thin enough to make the claim even approximately true will have, I suspect, very little to do with what we mean by the term.[6] (One is here not so far from—or at least on a slippery slope toward—the crude sociobiology that claims to observe "rape" in mallards or scorpionflies and concludes that rape must be "natural" for us.)[7] I would argue that all the interesting claims of this kind depend on just such oversimplistic uses of terms for complex social phenomena.
Third, and finally, I must take issue with the claims Konner makes about the genetic control of human behavior. At times, I must admit to being a little uncertain what these claims are. That there are "complex interactions of genotype, metabolism, and environment in the development of neural, behavioral, and mental phenomena" would be difficult to dispute. Yet I take it that the descriptions he offers of increasing scientific knowledge of genetically mediated mental disorders must be intended to show more than this. That these findings "remove any doubt regarding the ability of genes to alter complex behavior" suggests, if it does not strictly imply, that Konner has in mind the possibility of genetic control of complex behavior.[8]
At any rate, my objective here is not exegesis of Konner's essay. What does seem worth emphasizing is the utterly unsurprising character of the phenomena Konner draws to our attention and the absolute lack of any interesting conclusions about the determination of human behavior which follow from them. Like Konner, I think that "phenylketoneuria provides a valuable model." This, it will be recalled, is a genetic disorder that, in the absence of a specially controlled diet, results in massive toxicity to the brain and consequent psychological dysfunction. Apart from the banal fact that people with damaged or seriously abnormal brains are likely to behave strangely, it is very difficult to see what could follow from this. In particular, I cannot see that it tells us anything whatsoever about the relation between the genetically mediated development of the brain and mature human behavioral dispositions. To claim the contrary seems rather like concluding from the severe behavioral consequences of hitting someone over the head with a hammer that behavior must be determined to some important extent by an internal mechanism involving the action of miniature hammers. At the very least, not a shared of support is offered for hypotheses such as that there are genetic causes for, say, a predisposition to marriage.
Let me conclude on a more constructive note. I mentioned above that
there was more to an organism, especially a human one, than phylogeny and ontogeny. In the human case, I cannot state better, or phobably as well, what this is than Bernard Williams has done for us. The nature of a species, including our own, centrally includes its ethology; and the most striking and fundamental feature of human ethology is its existence without culture.
Without espousing some of the more excessive statements of cultural relativism, I think that the above fact requires that we take culture much more seriously than sociobiologists and their followers and apologists are generally prepared to do. A possible way of stating the consequences of so doing is the following.
We are incliding to suppose that because Homo sapiens is undoubtedly a perfectly respectable biological speices, its universal properties must provide the fundametnal insight into the nature and behavior of its members. But such taxonomic paralysis is just a form of essentialism, a traditional philosophical view almost uniformly rejected by contemporary theorists of biology.[9] If we reject essentialism, it is open to us to conclude, from the centrality of culture to human ethology and the great variability of culture, that for most purposes, Homo sapiens is much too broad and coarse a category for understanding human begings. We might more usefully think of humans not primarily as constituting one biological species but rather as composing many, no doubt overlapping, cultural species.[10]