Riparian Birds in the Riparian/Agricultural Interface[1]
Bertin W. Anderson, Robert D. Ohmart, and Hubert A. Allen, Jr.[2]
Abstract.—The interface between riparian and agricultural systems supports relatively large numbers of bird species and individuals because it offers a variety of food and structural resources that are especially apparent in winter. The interface can be used to compensate effectively for loss of natural habitats by interspersing agricultural lands with native vegetation.
Introduction
Conflict between a growing demand for more agricultural land and the need to maintain natural riparian systems is occurring all along the lower Colorado River. As valuable wildlife habitat continues to be converted to agricultural land, an urgent need exists to identify and preserve representative riparian systems and to determine how agricultural management might mitigate losses of natural habitats. This paper addresses the second point through a study of avian populations along the interface between riparian vegetation and cultivated lands.
We tested to see if the riparian/agricultural interface, hereafter referred to simply as "interface", is a distinct ecological community. Four avian community parameters were examined quantitatively in interface situations: 1) species richness (number of species); 2) avian densities; 3) species turnover (stability of species composition); and 4) relative proportions of different feeding guilds. Significant differences in these four parameters between agricultural land and riparian areas were used to indicate "distinctiveness." Finally we identify the unique attributes of the interface that define its value to riparian wildlife.
Methods
We censused each of eight riparian areas bordering agricultural cropland areas for two to three years between 1978 and 1980 using a modified Emlen technique (Emlen 1971, 1977; Anderson et al . 1977). Transects were located where relatively homogeneous stands of riparian vegetation adjoined agricultural fields. The interface transects had agricultural areas paralleling the transects on one side and riparian vegetation on the other. Two transects were located near Yuma, Arizona; two were on the Cibola National Wildlife Refuge near Palo Verde, California; and four were on the Colorado River Indian Reservation, between Ehrenberg and Parker, Arizona. Transects varied in length from 610 m. to 1,524 m.
Each transect was censused two or three times every month; results were averaged to obtain seasonal density estimates. Five seasons were considered: winter (December–February); spring (March–April); summer (May–July); late summer (August–September); and fall (October–November). These seasons reflect differences in the avian community as well as general climatic changes. The averaging of six to nine censuses minimized random variation inherent in the censusing method (Engel-Wilson etal . 1981).
Data for riparian vegetation were taken from Anderson and Ohmart (in preparation). Measurements in those vegetation-types of foliage density and diversity (MacArthur and MacArthur 1961), horizontal patchiness (Anderson etal . 1978), and tree and shrub densities were used to quantitatively characterize the vegetation of the transects. Crops in adjacent agricultural fields were noted when censuses were taken.
Bird species were classified as riparian if they occurred in riparian vegetation with some regularity (Conine et al . 1978). Riparian bird species were grouped into several feeding guilds based on general dietary habits. This grouping allowed us to examine the avian community in terms of a few functional components.
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Bertin W. Anderson is Faculty Research Associate, Center for Environmental Studies, Arizona State University, Tempe, Arizona. Robert D. Ohmart is Associate Director, Center for Environmental Studies, Arizona State University, Tempe, Arizona. Hubert A. Allen, Jr. is Field Biologist, Center for Environmental Studies, Arizona State University, Tempe, Arizona.
Changes in guild dominance within and between seasons indicated changes in community organization.
To calculate bird species turnover, a species list was compiled for each season by transect for each year. All riparian bird species were included except raptors, swallows, swifts, hummingbirds, very rare species, birds usually associated only with agricultural areas, and migrants. "Migrants" refers to those species which did not spend at least one of the five seasons; they were strictly transient in the area. These groups were excluded because they occurred rarely or inconsistently in riparian areas and their effects on the community were not clear. The resultant species list provided a conservative basis from which to calculate bird species turnover. A bird species "turned over" on a transect in a particular season if it was present one year and absent the next year, or vice versa. The species turnover rate was calculated as the mean number of turnovers per season divided by the mean number of species present per season.
The G-test and Mann-Whitney U-test were used in this analysis. These techniques were used to compare frequency ratios and nonparametric t-tests, respectively. Because the probability distribution function of avian species abundance is unknown, nonparametric analysis is recommended if values are to be compared (Siegel 1956; Sokal and Rolhf 1969). Mann-Whitney U-tests were used to compare numbers within each guild over the seasons. When using the Mann-Whitney U-test, two sets of abundance data were rank-ordered in a single array from highest to lowest. A test statistic was determined based on the ordering of the two samples. This statistic was compared to a theoretical distribution and a significance level was determined.
G-tests were used to test the goodness-off-it of frequency ratios. The hypothesis tested is that frequency ratios between replicate samples are homogeneous. We determined which season was consistently the richest by ranking seasonal richness values on each transect. Seasons then formed replicate samples. A five-part ratio of highest to second highest, etc., richness was compiled for the seasons, and a G-statistic was calculated. A significant value indicated that richness ratios were not equal among seasons.
Results
Riparian vegetation along the transects was composed primarily of salt cedar (Tamarix chinensis ), arrowweed (Tessariasericea ), and quail bush (Atriplexlentiformis ). Athel tamarisk (Tamarixaphylla ), a tall, evergreen species, was dominant at Yuma; salt cedar, a smaller deciduous species occurred in the other areas. Honey mesquite (Prosopisglandulosa ) and screwbean mesquite (Prosopispubescens ) trees were present on Cibola National Wildlife Refuge (CA) and Colorado River Indian Reservation (CRIR) transects. CRIR transects had high densities of shrubs other than arrowweed. Major crops in agricultural fields adjacent to transects included alfalfa, wheat, cotton, and pasture; in some seasons fields were fallow or plowed.
Bird species richness in interface was similar to that reported by Conine etal . (1978). Bird species richness was higher in interface than in either salt cedar vegetation or agricultural areas in all seasons except spring (fig. 1). Species numbers in cottonwood/willow (Populusfremontii/ Salixgooddingii ) vegetation, usually the richest riparian system, were higher than in interface in every season except winter, when the two systems had similar numbers of bird species. Honey mesquite vegetation averaged more bird species than interface in all seasons except late summer.
Bird species richness in interface varied significantly between seasons (P < 0.05). In interface, species richness values for late summer, fall, and winter were similar; they were higher than for spring and summer (fig. 1).
Interface bird densities were higher than those in either riparian or agricultural systems in all seasons except spring. Spring also averaged a higher turnover rate (32%) than other seasons in interface (fig. 2). Approximately 24% of bird species turned over between years (table 1). Except for spring, seasonal turnover rates were similar.
| ||||||||||||||||||||||||||||||||
Each avian guild showed distinct seasonal patterns of abundance (fig. 3). Numbers of resident insectivores were not significantly different from summer to winter, but spring densities were always lower (P < 0.05). Visiting insectivore densities showed a pattern similar to that of resident insectivores with significantly (P < 0.05) higher densities in fall and winter.
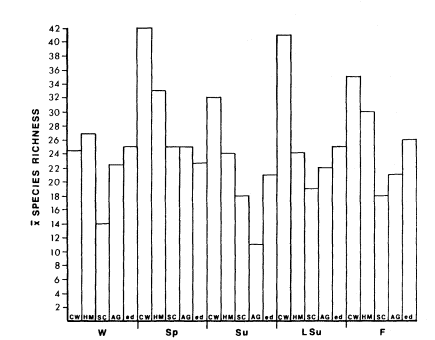
Figure l.
Average species richness in riparian and agricultural
(AG) systems of the lower Colorado River Valley.
CW = cottonwood/willow; HM = honey mesquite;
SC = salt cedar; ed = riparian/agricultural interface.
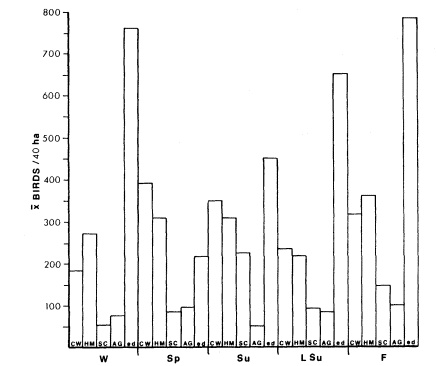
Figure 2.
Average densities in riparian and agricultural systems of the lower
Colorado River Valley. Abbreviations as in figure 1. W = winter;
Sp = spring; SU = summer; LSu = late summer; and F = fall.
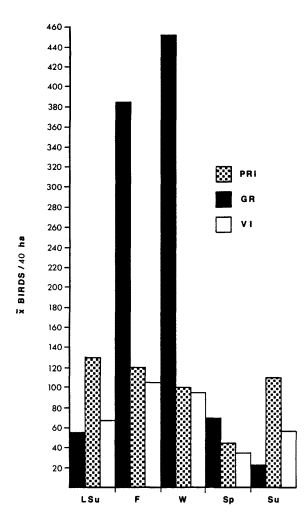
Figure 3.
Average densities in each feeding guild
in the riparian/agricultural interface. PRI
= permanent resident insectivores; VI =
visiting insectivores; GR = granivores.
Season abbreviations as in figure 2.
The relative proportion of individuals belonging to each feeding guild in interface also changed from season to season (fig. 4). Granivores were always proportionately dominant, but densities of species making up this group varied between seasons. The numbers of doves were highest in summer. Gambel Quail (Lophortyx gambelii ) numbers were highest in late summer. Other granivores, which were mostly sparrows, had peak densities during winter. The proportion of visiting insectivores was relatively stable, constituting about 15% of total individuals throughout the year. Resident insectivores were proportionately fewer in winter.
Discussion
Vegetation in the riparian/agricultural interface is often different from riparian vegetation in both plant species composition and structure. Interface areas tend to be subject to change, and as a result are more heterogeneous than other vegetation-types. Shrubs, especially quail bush and inkweed (Suaedatorreyana ), were more common in the interface than is usual in salt cedar stands. Quail bush shrubs have been shown to be related to increased bird densities in riparian systems (Anderson etal . 1978). These factors, as well as the juxtaposition of interface to agricultural fields, create a unique system.
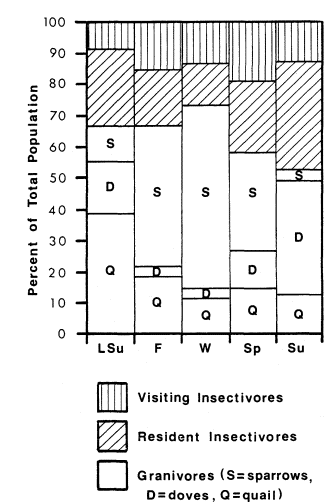
Figure 4.
Proportion of total population in the
riparian/agricultural interface belonging
to each feeding guild in each season.
Bird species richness and density comparisons indicate that when compared to salt cedar or agricultural systems, interface supported equal or greater numbers of birds in most seasons. Cottonwood/willow and honey mesquite vegetationtypes are generally considered the most diverse and productive in the lower Colorado River Valley; interface was similar in richness to honey mesquite, but compared favorably to cottonwood/willow only in winter. Bird densities in interface exceeded those in all communities in every season except spring.
The primary factor accounting for relative species richness of interface compared to salt cedar or agriculture is probably related to the diverse resources that are available there. Agricultural lands provide a nearly constant supply of food in the form of seeds, plants, and insects. However, they provide relatively poor cover and offer few nest or perch sites. Trees and dense shrubs in interface supply abundant cover and vertical structure for nesting and other activities. Many bird species that reach high densities in interface, including Mourning Dove (Zenaida macroura ), Gambel Quail, Whitecrowned Sparrow (Zonotrichialeucophrys ), and Loggerhead Shrike (Laniusludovicianus ), forage in agriculture but return to interface for protection from predators, nesting, and perching. This situation is an example of "interface effect"; the interface between two systems (riparian and agricultural) is richer than either system alone because elements of both are combined.
Most riparian systems show avian density peaks between summer and fall, but interface supported greater numbers of birds in fall and winter. This is partly because many individuals disperse out of their preferred riparian areas after the breeding season (Anderson and Ohmart in press). However, the winter peak in bird densities in interface is probably most closely related to food resources, which are relatively high in agricultural areas during winter due to year-round growing of crops, and irrigation. Food resources in riparian areas are lowest in winter (ibid .).
The dominance of granivores in interface in all seasons indicates that large supplies of seeds are available throughout the year. Seed sources include shrubs and annuals in interface, weedy margins around fields, and agricultural crops. A significant portion of the avian community in interface in all seasons is insectivores, which indicates that insect food resources are abundant throughout the year.
All three feeding guilds showed decreased use of interface at some time during the breeding season, indicating that it may not be preferred when resources are abundant in other areas or when other considerations, such as nest sites, are important.
Not all riparian bird species reach higher densities in interface than in riparian areas (Conine etal . 1978), but some species clearly benefit from association with interface. This is demonstrated by the Greater Roadrunner (Geococcyxcalifornianus ) and the Loggerhead Shrike. Both species are top-level carnivores that eat rodents, reptiles, and small birds. Both species generally breed in areas other than interface or agriculture, but they are present in interface from fall through winter. At these seasons, Loggerhead Shrikes and Greater Roadrunners reached significantly (P < 0.05) higher densities in interface than in riparian vegetation. These high densities indicate that interface is particularly rich in resources compared to other areas. Rice etal . (in press) determined that approximately one-third of all bird species turned over between years in riparian vegetation; this turnover was statistically random across habitats. Our method was more conservative than that of Rice etal . (ibid .) because we eliminated some bird species from consideration. Had we used the same method, species turnover rates in riparian and interface areas would have been similar. Thus, the avian composition of interface communities is no more stable than that of riparian communities. Species turnover rates of over 20% suggest that there is some opportunism in the use of interface. Some bird species may use interface only when conditions in preferred areas are limiting.
Conclusions
The interface between agricultural areas and riparian vegetation supports relatively large numbers of bird species and individuals, presumably because it offers a variety of food and structural resources. This is especially apparent in winter, when resources in riparian areas decline. Because interface accommodates large bird populations at a time when resources in other areas may be scarce, interface may be critical in maintaining avian populations that normally inhabit riparian areas or migrate into the lower Colorado River Valley.
Interface can be used effectively to mitigate loss of natural habitat components by interspersing agricultural lands with native vegetation. This will benefit many bird species which utilize food resources associated with agricultural areas but require cover or other features provided by natural vegetation. Interface could also be important in integrated pest management to help reduce pesticide use and costs in agriculture, although there are strong feelings opposing this idea. Maintenance of interface will not compensate for habitat loss for species that are riparian specialists, such as Summer Tanager (Pirangarubra ) or Yellow-billed Cuckoo (Coccyzusamericanus ). Protection of these species requires preservation or recreation of habitat in native riparian vegetation.
Acknowledgments
The authors wish to thank Marcelett Ector for typing various drafts of the manuscript and Cindy D. Zisner for typing the final draft. Susan M. Cook and Jane R. Durham provided editorial assistance. Elaine Hassinger prepared the figures. We are grateful to the many field biologists who assisted in collecting data. The project was funded by the USDI Bureau of Reclamation through Contract Number 7-07-30-V0009.
Literature Cited
Anderson, B.W., R.W. Engel-Wilson, D. Wells, and R.D. Ohmart. 1977. Ecological study of southwestern riparian habitats: techniques and data applicability. p. 146–155. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: a symposium. [Tuscon, Arizona, July 9, 1977]. USDA Forest Service GTR-RM-43. 217 p. Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colorado.
Anderson, B.W., and R.D. Ohmart. In preparation. Vegetation management. Annual report, 1981. USDI Bureau of Reclamation, Boulder City, Nevada.
Anderson, B.W., and R.D. Ohmart. In press. Evidence for social regulation in some riparian bird populations. Am. Nat. 120.
Anderson, B.W., R.D. Ohmart, and J. Disano. 1978. Revegetating the riparian floodplain for wildlife. p. 318–331. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: proceedings of the symposium. [Callaway Gardens, Georgia, December 11–13, 1978]. USDA Forest Service GTR-WO-12. 410 p. Washington, D.C.
Emlen, J.T. 1971. Population densities of birds derived from transect counts. Auk 88:323–342.
Emlen, J.T. 1977. Estimating breeding season bird densities from transect counts. Auk 94:455–468.
Engel-Wilson, R.W., A.K. Webb, K.V. Rosenberg, R.D. Ohmart, and B.W. Anderson. 1981. Avian censusing with the strip method: a computer simulation. p. 445–449. In : C.J Ralph and J.M. Scott (ed.). Estimating numbers of terrestrial birds. [Asilomar, California, October 26–31, 1980]. Studies in Avian Biology No. 6.
MacArthur, R.H., and J.W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.
Rice, J.C., R.D. Ohmart, and B.W. Anderson. In press. Rates of change in species composition of avian communities in contiguous riparian habitats. Ecology.
Siegel, S. 1956. Nonparametric statistics for the behavioral sciences. 312 p. McGraw-Hill, New York, N.Y.
Sokal, R.R., and R.J. Rolhf. 1969. Biometry. 776 p. W.H. Freeman and Company, San Francisco, California.