Paleozoic And Mesozoic Gymnosperms
(Arnold 1969; Axelrod 1974; Bambach et al. 1980; Barnard 1973; Beck 1976; Brenner 1976; Chaloner and Lacey 1973; Chaloner and Meyen 1973; Douglas and Williams 1982; Florin 1963; Hallam 1977; Hughes 1976; Mamay 1976; Miller 1977; Penny 1969; Phillips et al. 1985; Plumstead 1973; Rabinowitz et al. 1983; Retallack and Dilcher 1981a ; Roche 1974; Stewart 1983; Taylor 1981)
Origin of Gymnosperms
About 3 billion years passed between the appearance in the fossil record of the first plants with chlorophyll, the cyanobacteria or blue-green algae, and the appearance of the first vascular land plants, the Silurian pteridophytes.
These non-seed-bearing pteridophytes diversified during the Devonian into several major lineages, commonly interpreted as having surviving members in the whiskferns, horsetails, clubmosses, and ferns. All these evolved complex vascular diploid bodies, which in some cases were large trees. Once established, a diploid pteridophyte is well fitted for survival on land. Also, the microscopic haploid spores, which pteridophytes produce in huge quantities, are well adapted to long-range wind dispersal. After the spores land on the ground and germinate, however, the ensuing haploid stages, including a swimming sperm, are not much better fitted for land life than their algal ancestors.
Evolution by the gymnosperms of pollen and seeds eliminated the weak links in the pteridophyte life cycle for sexual reproduction on land. The evolutionary steps were complex, but the net result was simply sheltering and nurturing the vulnerable stages within the body tissues of the mature vascular diploid. Gymnosperm cones are organs of the diploid plant and are supplied with water and food by its vascular tissues. The only stage of the sexual reproductive cycle that has to exist out in the world on its own is the haploid pollen. The target of a pollen grain, unlike a fern spore, is not the ground but an opening in the integument surrounding the young seed on a female cone. There it can deliver the sperm to the egg without external water.
Origin of the gymnosperms came remarkably soon after the origin of their pteridophyte ancestors. An intermediate group, the progymnosperms, appeared in the Devonian in what is now North America and Europe. The geography of the Paleozoic world is still very speculative, but reconstructions for the Devonian and early Carboniferous commonly show North America, Greenland, and Europe united as a single continent, called Euramerica, which was located in low latitudes and separated by oceans from proto-Asia and Gondwanaland. The oceans had evidently not barred pteridophyte migrations; from the Silurian to the Carboniferous, pteridophyte floras had fairly uniform global distributions. The seeds of early gymnosperms, however, like those of most modern ones, were evidently poorly equipped for long-range wind or water dispersal. Although gymnosperm pollen grains are capable of long-range wind dispersal, this serves only to maintain gene flow between already established disjunct populations of a species, not to found a new population.
So far as is known at present, the first gymnosperms were confined to Euramerica. The first named seed plant species, Archaeosperma arnoldii , appeared there in Late Devonian time. Its seeds were borne in pairs on open, cuplike structures arranged more loosely than in the tightly condensed cones of later gymnosperms, and there was a larger opening in the integument for entry of the pollen than in later gymnosperms.
Carboniferous
The Carboniferous is named for the remains of the great swamp forests laid down as coal. In both Europe and North America, the abundant fossil record shows that these swamps were dominated by pteridophyte trees, with gymnosperms as subordinate associates. The nature of the upland flora, if any, is not known.
The gymnosperms of the Early Carboniferous (or Mississippian) already represented several families within the extinct order of Pteridospermales or seed ferns. Two other gymnosperm orders, presumably also derived from the progymnosperms, namely the Cordaitales and Voltziales, became widespread in both midcontinental North America and western Europe in the late Carboniferous (Pennsylvanian). Both were structurally closer to modern conifers than to ferns.
Some paleogeographic reconstructions show Pangea beginning to be assembled in the Late Carboniferous, with proto-Asia and Gondwanaland approaching or contacting Euramerica. This would account for reports of Pennsylvanian age gymnosperms of all three orders in parts of Asia and Gondwanaland, although dating and identity of these fossils is not very clear. Even if Pangea was assembled, there were great barriers to seed plant migration during the Pennsylvanian because of extensive marine transgressions and, in Gondwanaland, extensive glaciation.
Permian
During Permian time, Pangea was complete, with broad contacts near the equator between Euramerica and western Gondwanaland. A single world ocean extended from pole to pole and spanned 300° of longitude at the equator. The interior of the continent must have had great deserts, particularly behind upthrust mountains; shallow inland seas created other barriers to seed plant migration. In spite of contacts between continental plates, the Permian had regional floristic differences.
The three Carboniferous gymnosperm orders, Pteridospermales, Cordaitales, and Voltziales, survived in the Permian in Euramerica and were joined by probable primitive representatives of two living gymnosperm orders, Cycadales and Ginkgoales. During the Permian, all five lineages appeared in Asia and Gondwanaland, where they were mainly represented by different genera than in Euramerica. The Cordaitales became extinct late in the Permian.
The most famous Gondwana gymnosperm genus is Glossopteris , which
can be classified either as a member of the Pteridospermales or in an order of its own. Glossopteris was a tree with trunks as large as 40 cm in diameter and with narrow, tongue-shaped leaves. It dominated coastal swamps that accumulated to form much of the commercial coal of the southern hemisphere, uplands being occupied by other gymnosperms. In the Permian, Glossopteris spread over huge areas of present South America, Africa, India, Antarctica, and Australia that had been glaciated during the Carboniferous. The paleolatitudes within which Glossopteris grew ranged from about 40°S to almost the Permian south pole (fig. 13). It is not known whether the trees were evergreen or deciduous, but in any case, it is hard to account for their survival in nearly six months without sun.
Triassic and Jurassic
The Mesozoic began with Pangea fully assembled and with low sea levels. During Triassic time, the Tethys Sea formed between the single northern continent, Laurasia, and Gondwanaland, but it remained narrow in places; there may have been intermittent contacts between the northern and southern continents during much of the Mesozoic. By the Middle Jurassic, sea level had risen and shallow continental seas separated North America—Greenland from Europe and South America from Africa, at least intermittently. Also during the Jurassic, India-Madagascar may have already broken away from Africa.
Glossopteris survived through the Triassic but died out early in the Jurassic, ending one pteridosperm lineage. Two other now extinct orders possibly derived from the pteridosperms, namely the Caytoniales and the Bennettitales, arose during the Triassic. The Caytoniales had a fleshy tissue partly enclosing the seed that suggests animal dispersal. One genus of Caytoniales, Lepidopteris , had a remarkable range across Laurasia and Gondwanaland in paleolatitudes from 60°N to 60°S. The Bennettitales, which superficially resemble cycads, had spectacular reproductive structures that suggest adaptation for animal pollination.
The two Paleozoic gymnosperm orders that survive today, Cycadales and Ginkgoales, spread widely through both Laurasia and Gondwanaland in the Mesozoic, increasing greatly in abundance and diversity. Some Jurassic Ginkgo leaves are indistinguishable from those of the living G. biloba, and some of the cycads were already similar to living genera.
Two other living orders of gymnosperms, Coniferales and Taxales, arose in the Mesozoic not long before the Voltziales, their probable progenitors, became extinct. Five important modern conifer families had evidently already diverged by the Early Jurassic: Pinaceae in Laurasia, Podocarpaceae in Gondwanaland, and Araucariaceae, Cupressaceae, and Taxodiaceae on both
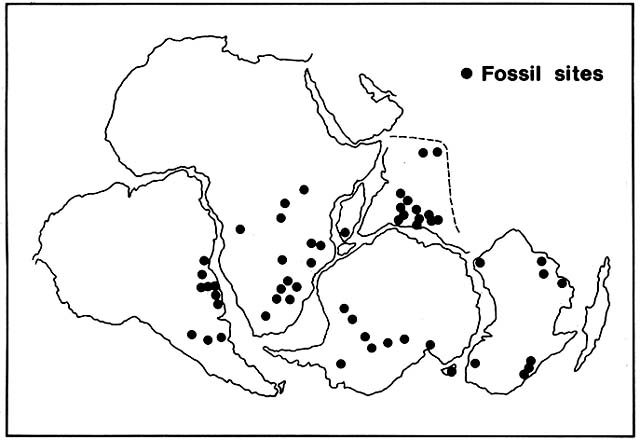
Figure 13. Late Paleozoic Range of Glossopteris , a Dominant Seedfern on the Supercontinent of Gondwanaland.
Gondwanaland broke up into several continents, outlined by dashed lines. Fossils of Glossopteris were then
displaced to widely separated parts of the globe. Since the genus evidently was not adapted for transoceanic
dispersal, its fossil distribution appears inexplicable on an earth with fixed continents. This was noted as evidence
for the hypothesis of continental drift long before the modern theory of seafloor spreading was developed.
continents. During the Jurassic, the Taxales, precursors of the living yew, entered the fossil record in Laurasia.
Cretaceous
The Tethys Sea continued to be a partial barrier between the northern and southern continents during Cretaceous time, but there were intermittent contacts between Europe and Africa and between North and South America. During the Early Cretaceous, the incipient Indian and South Atlantic oceans began to dissect Gondwanaland, but separation was incomplete except for isolation of India–Madagascar during most of the Cretaceous. By the Santonian, Africa was an island, but South America may still have been linked via Antarctica with Australia and New Zealand. By the Maestrichtian, the South Atlantic had widened to about 800 km between Brazil and Africa, but there had been little seafloor spreading in the North Atlantic. North America, Greenland, and Europe were still in contact north of 50°N latitude. At higher latitudes, Alaska and Asia had been in contact since the Coniacian. Laurasia, however, was divided during much of the Cretaceous by marine transgressions. Although the extensive Jurassic epicontinental seas had receded at the start of the Cretaceous, the regression was temporary. Seaways extending from the Tethys Sea to the Arctic Ocean bisected both North America and Eurasia from mid-Cretaceous until they withdrew in the Maestrichtian.
Two Triassic gymnosperm orders; the Caytoniales and Bennettitales, remained widespread in the Early Cretaceous but were extinct by the end of the Cretaceous.
Cycadales and Ginkgoales continued to be widespread throughout the Cretaceous, ranging to amazingly high latitudes in both northern and southern continents. The Taxales remained confined to Laurasia.
Among the Coniferales, the Pinaceae were represented by fossils confined to Laurasia; extant genera represented included Pinus and possibly Cedrus and Larix . The Podocarpaceae continued to be widespread in parts of former Gondwanaland, excluding Africa; fossils are commonly identified with living genera, especially Dacrydium and Podocarpus . The Araucariaceae, Cupressaceae, and Taxodiaceae continued their Jurassic pattern of wide ranges in both northern and southern continents. Jefferson (1982) described a puzzling fossil forest of conifers resembling Araucaria that lived during the Early Cretaceous in Antarctica at a paleolatitude of about 70°S. The trees were buried by volcaniclastic deposits and remained standing and rooted. The trees must have had problems of shading under the low sun angle, but they were closely spaced and had an understory of pteridophyte vegetation. Also, their well-defined growth rings show that annual growth was at a rate that would be very high for modern conifers, even at low latitudes.
Among the Taxodiaceae, fossils tentatively assigned to Sequoiadendron , which survives only in California, were deposited in eastern North America and Greenland in Early Cretaceous time (Florin 1963). By Late Cretaceous, this taxon ranged to Europe and western North America. Metasequoia , which survives only in China, had a coherent Cretaceous range from eastern Asia through Alaska to western North America. Sequoia , now confined to western North America, evidently originated there, perhaps by hybridization between Sequoiadendron and Metasequoia . Various other genera of Taxodiaceae had close Cretaceous precursors. Cryptomeria, Cunninghamia, Glyptostrobus , and Sciadopitys , which survive only in eastern Asia, had Cretaceous precursors in widely scattered sites in Eurasia, North America, and Greenland. Arthrotaxis , which survives only in Tasmania, had Cretaceous precursors in New Zealand and South America.