1.9—
The Role of the Plant Cell Wall in Interactions with Other Organisms
Plants are beset by many pests and pathogens and helped by a variety of symbionts. The walls of epidermal cells may be specialized in a number of ways to form a protective skin over the entire plant (e.g. cutin, wax, gums and mucilages, bark and thorns, etc.). Many cell walls throughout the plant can become resistant to degradation by pathogens by means of lignification.
Quite apart from such modifications, however, the structure of the cell wall
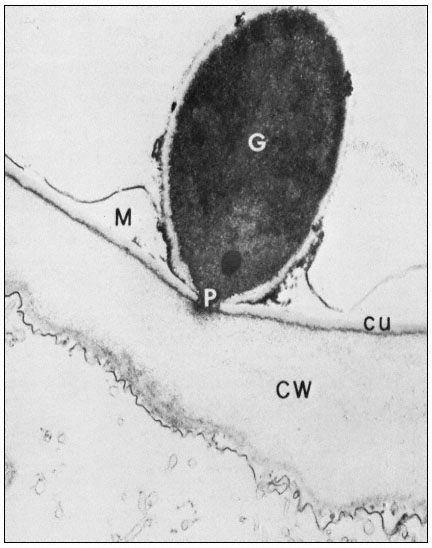
Figure 1.10
Penetration of epidermal cell walls of broad bean by Botrytis cinerea.
The fungal germ tube (G) is attached to the surface of the cuticle (CU) by
a layer of mucilage (M) secreted by the fungus. The fungus appears to
have dissolved a hole through the cuticle and to have begun dissolving
the plant cell wall (CW) beneath this hole. A membrane-bound infection
peg (P) penetrates through the pore in the fungal cell wall and the cuticle.
(Courtesy Prof. W. E. McKeen. See Phytopathology (1974), 64, 461–67.)
can itself present a formidable barrier to pathogens. In order to penetrate the cell wall and utilize its component sugars, pathogenic fungi and bacteria have evolved sophisticated, inducible batteries of enzymes which can hydrolyze components of the plant cell wall (Bateman & Basham, 1975). Figure 1.10 illustrates the initial stages of cell wall penetration by a fungal pathogen. The pathogen appears to have dissolved a hole through the cuticle and to have started degrading the cell wall.
Plants, in turn, have evolved countermeasures to the hydrolytic enzymes of the pathogens. In primary walls, the molecular architecture is such that only the pectic polysaccharides are accessible to hydrolytic enzymes (Bauer et al., 1973). It is probably for this reason that the first enzymes to be secreted by an invading pathogen are the pectin-hydrolyzing enzymes (Bateman & Basham, 1975). This is also likely to be the reason why the walls of many plants contain proteins which specifically inhibit the pectin-degrading enzymes (and only the pectin-degrading enzymes) of pathogenic microorganisms (Anderson & Albersheim, 1971). In other plants the pectic polysaccharides are heavily acetylated, and thus resistant to enzymic attack.
Plants have evolved mechanisms for counterattack as well as defence. Inducible enzymes are present in the cell walls of several plants which hydrolyze the wall polysaccharides of invading fungal pathogens (Ables et al., 1970; Pegg & Vessey, 1973). The pathogens react by secreting proteins which specifically inhibit the attacking plant enzymes (Albersheim & Valent, 1974). The cell walls of a variety of other plants contain glycoside-hydrolyzing enzymes which can release hydrogen cyanide from cyanogenic glycosides. The release of hydrogen cyanide by the plant occurs in response to attack by a pathogen. The pathogen (sometimes) avoids cyanide poisoning by an inducible enzyme which converts the cyanide to harmless formamide (Fry & Munch, 1975).
From these and other examples it is clear that the cell wall is a most important battleground in the contest between plants and their pathogens. The plant cell wall is not just a strong but passive barrier to invasion. It is impregnated with a host of molecules which can recognize a pathogen, modify the defences, or mount a counterattack.