9
The Deep Past
The Neogene and later periods discussed above cover less than one-tenth of the time that seed plants have existed. Their fossil record, which has great gaps in space and time, began in the Devonian Period (Table 7), more than 300 million years before the Neogene. Unlike the Neogene, for which nearly all fossil plants can be identified to living genera, if not species, the of characters becomes increasingly strange with greater age. More often than not, it is highly speculative whether these strange creatures were evolutionary dead ends or primitive ancestors of much changed modern descendants. Moreover, paleontologists not only have to deal with strange creatures, they usually have only bits of creatures that nobody has seen whole. Separate taxonomies have to be constructed for pollen, leaves, stems, seeds, and other organs. Articulating them is a great accomplishment. For example, the existence of a great extinct order, the bizarre seed ferns or pteridosperms, was not revealed until it was finally concluded that fossil seeds, long named Lagenostoma, and fernlike foliage, named Lyginopteris, belonged to the same plant.
Also, research on early seed plants has necessarily been partitioned among specialists in different taxonomic groups whose first task was to work out evolutionary relationships of particular lineages. Only gradually have plant geographic syntheses emerged.
For this time span, another change in approach to migrations is necessary. There is no hope of tracing geographic changes on the species level. Even for higher taxa, information on geographic distributions is generally rudimentary. All that can be attempted here is to seek evidence of really grand migrations of higher taxa. This will be done chronologically, starting with the origins of the first seed plants. Some notice will be taken of continental movements, but for all of the dazzling light that plate tectonics has shed on
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table continued on next page
| ||||||||||||||||||||||||||||||||||||||||||||||||
geomorphology, it is still of limited use in reconstructing plant migrations. Contact between tectonic plates is irrelevant if the continental margins were submerged. A shallow sea is as much a barrier for land plants incapable of long-range dispersal as is a deep ocean.
Paleozoic And Mesozoic Gymnosperms
(Arnold 1969; Axelrod 1974; Bambach et al. 1980; Barnard 1973; Beck 1976; Brenner 1976; Chaloner and Lacey 1973; Chaloner and Meyen 1973; Douglas and Williams 1982; Florin 1963; Hallam 1977; Hughes 1976; Mamay 1976; Miller 1977; Penny 1969; Phillips et al. 1985; Plumstead 1973; Rabinowitz et al. 1983; Retallack and Dilcher 1981a ; Roche 1974; Stewart 1983; Taylor 1981)
Origin of Gymnosperms
About 3 billion years passed between the appearance in the fossil record of the first plants with chlorophyll, the cyanobacteria or blue-green algae, and the appearance of the first vascular land plants, the Silurian pteridophytes.
These non-seed-bearing pteridophytes diversified during the Devonian into several major lineages, commonly interpreted as having surviving members in the whiskferns, horsetails, clubmosses, and ferns. All these evolved complex vascular diploid bodies, which in some cases were large trees. Once established, a diploid pteridophyte is well fitted for survival on land. Also, the microscopic haploid spores, which pteridophytes produce in huge quantities, are well adapted to long-range wind dispersal. After the spores land on the ground and germinate, however, the ensuing haploid stages, including a swimming sperm, are not much better fitted for land life than their algal ancestors.
Evolution by the gymnosperms of pollen and seeds eliminated the weak links in the pteridophyte life cycle for sexual reproduction on land. The evolutionary steps were complex, but the net result was simply sheltering and nurturing the vulnerable stages within the body tissues of the mature vascular diploid. Gymnosperm cones are organs of the diploid plant and are supplied with water and food by its vascular tissues. The only stage of the sexual reproductive cycle that has to exist out in the world on its own is the haploid pollen. The target of a pollen grain, unlike a fern spore, is not the ground but an opening in the integument surrounding the young seed on a female cone. There it can deliver the sperm to the egg without external water.
Origin of the gymnosperms came remarkably soon after the origin of their pteridophyte ancestors. An intermediate group, the progymnosperms, appeared in the Devonian in what is now North America and Europe. The geography of the Paleozoic world is still very speculative, but reconstructions for the Devonian and early Carboniferous commonly show North America, Greenland, and Europe united as a single continent, called Euramerica, which was located in low latitudes and separated by oceans from proto-Asia and Gondwanaland. The oceans had evidently not barred pteridophyte migrations; from the Silurian to the Carboniferous, pteridophyte floras had fairly uniform global distributions. The seeds of early gymnosperms, however, like those of most modern ones, were evidently poorly equipped for long-range wind or water dispersal. Although gymnosperm pollen grains are capable of long-range wind dispersal, this serves only to maintain gene flow between already established disjunct populations of a species, not to found a new population.
So far as is known at present, the first gymnosperms were confined to Euramerica. The first named seed plant species, Archaeosperma arnoldii , appeared there in Late Devonian time. Its seeds were borne in pairs on open, cuplike structures arranged more loosely than in the tightly condensed cones of later gymnosperms, and there was a larger opening in the integument for entry of the pollen than in later gymnosperms.
Carboniferous
The Carboniferous is named for the remains of the great swamp forests laid down as coal. In both Europe and North America, the abundant fossil record shows that these swamps were dominated by pteridophyte trees, with gymnosperms as subordinate associates. The nature of the upland flora, if any, is not known.
The gymnosperms of the Early Carboniferous (or Mississippian) already represented several families within the extinct order of Pteridospermales or seed ferns. Two other gymnosperm orders, presumably also derived from the progymnosperms, namely the Cordaitales and Voltziales, became widespread in both midcontinental North America and western Europe in the late Carboniferous (Pennsylvanian). Both were structurally closer to modern conifers than to ferns.
Some paleogeographic reconstructions show Pangea beginning to be assembled in the Late Carboniferous, with proto-Asia and Gondwanaland approaching or contacting Euramerica. This would account for reports of Pennsylvanian age gymnosperms of all three orders in parts of Asia and Gondwanaland, although dating and identity of these fossils is not very clear. Even if Pangea was assembled, there were great barriers to seed plant migration during the Pennsylvanian because of extensive marine transgressions and, in Gondwanaland, extensive glaciation.
Permian
During Permian time, Pangea was complete, with broad contacts near the equator between Euramerica and western Gondwanaland. A single world ocean extended from pole to pole and spanned 300° of longitude at the equator. The interior of the continent must have had great deserts, particularly behind upthrust mountains; shallow inland seas created other barriers to seed plant migration. In spite of contacts between continental plates, the Permian had regional floristic differences.
The three Carboniferous gymnosperm orders, Pteridospermales, Cordaitales, and Voltziales, survived in the Permian in Euramerica and were joined by probable primitive representatives of two living gymnosperm orders, Cycadales and Ginkgoales. During the Permian, all five lineages appeared in Asia and Gondwanaland, where they were mainly represented by different genera than in Euramerica. The Cordaitales became extinct late in the Permian.
The most famous Gondwana gymnosperm genus is Glossopteris , which
can be classified either as a member of the Pteridospermales or in an order of its own. Glossopteris was a tree with trunks as large as 40 cm in diameter and with narrow, tongue-shaped leaves. It dominated coastal swamps that accumulated to form much of the commercial coal of the southern hemisphere, uplands being occupied by other gymnosperms. In the Permian, Glossopteris spread over huge areas of present South America, Africa, India, Antarctica, and Australia that had been glaciated during the Carboniferous. The paleolatitudes within which Glossopteris grew ranged from about 40°S to almost the Permian south pole (fig. 13). It is not known whether the trees were evergreen or deciduous, but in any case, it is hard to account for their survival in nearly six months without sun.
Triassic and Jurassic
The Mesozoic began with Pangea fully assembled and with low sea levels. During Triassic time, the Tethys Sea formed between the single northern continent, Laurasia, and Gondwanaland, but it remained narrow in places; there may have been intermittent contacts between the northern and southern continents during much of the Mesozoic. By the Middle Jurassic, sea level had risen and shallow continental seas separated North America—Greenland from Europe and South America from Africa, at least intermittently. Also during the Jurassic, India-Madagascar may have already broken away from Africa.
Glossopteris survived through the Triassic but died out early in the Jurassic, ending one pteridosperm lineage. Two other now extinct orders possibly derived from the pteridosperms, namely the Caytoniales and the Bennettitales, arose during the Triassic. The Caytoniales had a fleshy tissue partly enclosing the seed that suggests animal dispersal. One genus of Caytoniales, Lepidopteris , had a remarkable range across Laurasia and Gondwanaland in paleolatitudes from 60°N to 60°S. The Bennettitales, which superficially resemble cycads, had spectacular reproductive structures that suggest adaptation for animal pollination.
The two Paleozoic gymnosperm orders that survive today, Cycadales and Ginkgoales, spread widely through both Laurasia and Gondwanaland in the Mesozoic, increasing greatly in abundance and diversity. Some Jurassic Ginkgo leaves are indistinguishable from those of the living G. biloba, and some of the cycads were already similar to living genera.
Two other living orders of gymnosperms, Coniferales and Taxales, arose in the Mesozoic not long before the Voltziales, their probable progenitors, became extinct. Five important modern conifer families had evidently already diverged by the Early Jurassic: Pinaceae in Laurasia, Podocarpaceae in Gondwanaland, and Araucariaceae, Cupressaceae, and Taxodiaceae on both
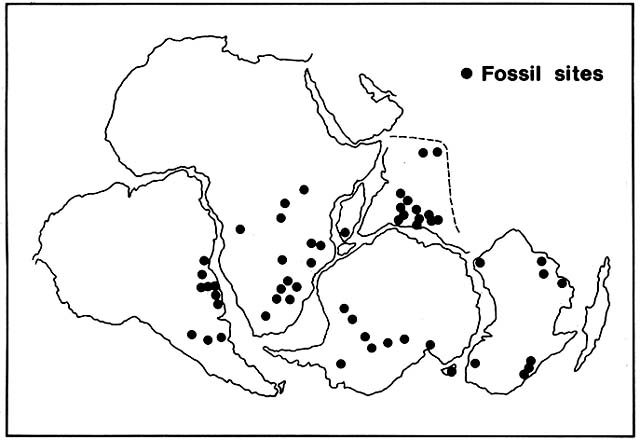
Figure 13. Late Paleozoic Range of Glossopteris , a Dominant Seedfern on the Supercontinent of Gondwanaland.
Gondwanaland broke up into several continents, outlined by dashed lines. Fossils of Glossopteris were then
displaced to widely separated parts of the globe. Since the genus evidently was not adapted for transoceanic
dispersal, its fossil distribution appears inexplicable on an earth with fixed continents. This was noted as evidence
for the hypothesis of continental drift long before the modern theory of seafloor spreading was developed.
continents. During the Jurassic, the Taxales, precursors of the living yew, entered the fossil record in Laurasia.
Cretaceous
The Tethys Sea continued to be a partial barrier between the northern and southern continents during Cretaceous time, but there were intermittent contacts between Europe and Africa and between North and South America. During the Early Cretaceous, the incipient Indian and South Atlantic oceans began to dissect Gondwanaland, but separation was incomplete except for isolation of India–Madagascar during most of the Cretaceous. By the Santonian, Africa was an island, but South America may still have been linked via Antarctica with Australia and New Zealand. By the Maestrichtian, the South Atlantic had widened to about 800 km between Brazil and Africa, but there had been little seafloor spreading in the North Atlantic. North America, Greenland, and Europe were still in contact north of 50°N latitude. At higher latitudes, Alaska and Asia had been in contact since the Coniacian. Laurasia, however, was divided during much of the Cretaceous by marine transgressions. Although the extensive Jurassic epicontinental seas had receded at the start of the Cretaceous, the regression was temporary. Seaways extending from the Tethys Sea to the Arctic Ocean bisected both North America and Eurasia from mid-Cretaceous until they withdrew in the Maestrichtian.
Two Triassic gymnosperm orders; the Caytoniales and Bennettitales, remained widespread in the Early Cretaceous but were extinct by the end of the Cretaceous.
Cycadales and Ginkgoales continued to be widespread throughout the Cretaceous, ranging to amazingly high latitudes in both northern and southern continents. The Taxales remained confined to Laurasia.
Among the Coniferales, the Pinaceae were represented by fossils confined to Laurasia; extant genera represented included Pinus and possibly Cedrus and Larix . The Podocarpaceae continued to be widespread in parts of former Gondwanaland, excluding Africa; fossils are commonly identified with living genera, especially Dacrydium and Podocarpus . The Araucariaceae, Cupressaceae, and Taxodiaceae continued their Jurassic pattern of wide ranges in both northern and southern continents. Jefferson (1982) described a puzzling fossil forest of conifers resembling Araucaria that lived during the Early Cretaceous in Antarctica at a paleolatitude of about 70°S. The trees were buried by volcaniclastic deposits and remained standing and rooted. The trees must have had problems of shading under the low sun angle, but they were closely spaced and had an understory of pteridophyte vegetation. Also, their well-defined growth rings show that annual growth was at a rate that would be very high for modern conifers, even at low latitudes.
Among the Taxodiaceae, fossils tentatively assigned to Sequoiadendron , which survives only in California, were deposited in eastern North America and Greenland in Early Cretaceous time (Florin 1963). By Late Cretaceous, this taxon ranged to Europe and western North America. Metasequoia , which survives only in China, had a coherent Cretaceous range from eastern Asia through Alaska to western North America. Sequoia , now confined to western North America, evidently originated there, perhaps by hybridization between Sequoiadendron and Metasequoia . Various other genera of Taxodiaceae had close Cretaceous precursors. Cryptomeria, Cunninghamia, Glyptostrobus , and Sciadopitys , which survive only in eastern Asia, had Cretaceous precursors in widely scattered sites in Eurasia, North America, and Greenland. Arthrotaxis , which survives only in Tasmania, had Cretaceous precursors in New Zealand and South America.
Cretaceous Angiosperms
(Axelrod and Raven 1978; Batten 1981; Beck 1976; Brenner 1976; Dilcher and Crane 1984, 1985; Doyle 1973, 1977; Doyle and Hickey 1976; Hickey 1977; Hughes 1976; Muller 1970, 1981; Retallack and Dilcher 1981a,b ; Srivastava 1981; Tralau 1973; Tschudy and Scott 1969; Walker and Doyle 1975; Walker et al. 1983)
The origin of the angiosperms has long been one of the most baffling problems in paleobotany. Various fossil gymnosperm orders and all living ones except the conifers have been suggested as possible progenitors, but missing links in the fossil record make all the alternatives highly speculative. Until recently it was generally supposed that the angiosperms originated someplace where there was no fossil record and underwent major phylogenetic diversification, including initiation of many living families, before entering the fossil record. Major theories of angiosperm phylogeny have been built almost entirely by deductive comparisons between living groups, with any relevant fossil evidence considered to be simply missing.
Recently, paleobotanists have launched a new attack on the problem of Cretaceous angiosperm evolution, and hopes are currently high that the fossil record is much more complete than had been realized. As noted by Wolfe (1973), the amount of missing record may have been greatly exaggerated by a practice by pioneer paleobotanists of seeking resemblances between Cretaceous leaf fossils and living genera and assigning the fossil to whatever modern family offered the closest resemblance. This practice has worked well for Neogene and Pleistocene woody species that have undergone very little subsequent evolution or extinction. In these relatively young fossil floras,
identifications based on leaves, other macrofossils, and pollen have all fit together nicely. Identifications of Cretaceous leaves with modern taxa, however, have not generally been supported by other fossils, and resemblances may be merely superficial.
Instead of leaping to identify primitive angiosperm fossils with familiar names, efforts are now being made to work forward from the beginning of the record in early Cretaceous, to build an independent taxonomy of initially strange pollen and macrofossils, which are not expected to have already become stabilized in their present forms. Pollen is being given special attention because it provides a more abundant and complete sampling of the flora from a catchment area larger than that of the macrofossils. By comparative study of enough floras worked up in this way, it may eventually become possible to trace angiosperm evolution and migration through the Cretaceous and to follow some lineages to extinction and others to recognizable modern descendants. Only glimpses of such a story are now possible.
Neocomian
The earliest generally accepted fossil evidence of an angiosperm comes from beds deposited over 100 million years ago in Siberia; the fossil consists of small, pinnately veined, simple leaves, presumably from a dicot. How far this plant had proceeded in transition from gymnosperm to angiosperm reproductive structures is unknown.
Barremian
A sparse macrofossil record of apparent monocot and dicot leaves and of dicot fruits and a more abundant pollen record suggesting both monocots and dicots are reported from both sides of the Tethys Sea in North America, Europe, Siberia, South America, and Africa. The angiosperm fossils were deposited in sediments indicating that they inhabited shores of lagoons, estuaries, and other sites disturbed by coastal erosion and deposition. Retallack and Dilcher (1981b ) suggested that the angiosperms originated as coastal pioneers capable of rapid migration by wind and water dispersal. The ocean may have served more as a highway than as a barrier to migration, as is true of later seashore angiosperms, which typically have seeds or fruits dispersed by ocean currents. Because of the pervasive symbioses between angiosperms and animal pollinators and seed dispersers, it has been widely assumed that the very origin of angiospermy involved coevolution with animals, for example, that the ovary served initially to protect the young ovules from chewing insects visiting the flowers. If the angiosperms really arose as coastal pioneers,
perhaps the ovary served initially for sea dispersal and coevolution with animals was secondary.
Barremian pollen attributed to angiosperms include a probable monocot form, Liliacidites , and a probable dicot form, Clavatipollenites . Both resemble gymnosperm pollen in being monosulcate, that is, having a single germination furrow, but differ from gymnosperm pollen in having intricately sculptured instead of smooth coats. This sculpturing may have adapted the pollen grain to adhere to the body of an insect or to the stigma of a flower. The latter would mean that the plants had crossed the crucial line defining angiospermy, that is, the ovules were no longer exposed to direct pollination but had to be reached by growth of a long pollen tube penetrating the ovary from a stigma. Among living angiosperms similar primitive monosulcate pollen is produced by many monocots and by one of the six subclasses of dicots, namely, the Magnoliidae, which include Magnoliaceae, Lauraceae, Nymphaeaceae, and related families.
Aptian
Monosulcate angiosperm pollen continued to be widespread in both the northern and southern hemispheres. In Gondwanaland, it was joined by the first tricolpate pollen, that is, grains with three germination furrows, almost certain proof of angiospermy. In the moist pollen chamber of a gymnosperm ovule, a single germination locus suffices, but on the exposed stigma of an angiosperm flower, multiple pores on different sides of the grain are highly advantageous. Aptian tricolpate pollen has been found in Israel in sediments deposited at low latitudes on the Gondwanaland coast and also in sediments deposited along embayments in the rift valley system lying between what is now equatorial Africa and South America. Tricolpate pollen and forms with similar names derived from it by elaboration and modification are characteristic of all the six subclasses of dicots except the Magnoliidae.
Albian
During the Albian, tricolpate and derivative pollen types diversified vigorously, not only in Gondwanaland but also north of the Tethys Sea in both western and eastern North America and in Europe. Later in the Albian, angiosperms evidently spread to higher latitudes than ever before in both Laurasia and Gondwanaland; this expansion is indicated by both monosulcate and tricolpate pollen and by macrofossils. In North America and Europe, dicot leaves became much more diverse; in addition to forms that suggest Magnoliidae, leaves appeared that may represent other dicot subclasses, such
as Hamamelidae, exemplified by Platanus . Actual fossilized flowers are known from the Albian of North America and Europe; they indicate divergent adaptations for insect and wind pollination. Some are bisexual, with large, showy petals and many ovules, closely resembling Magnolia . Some are bisexual and have small, delicate petals, probable nectaries, and few ovules, closely resembling Pyrus , the apple. Some are unisexual, tiny, inconspicuous, and grouped in inflorescences strikingly similar to Platanus .
Generally angiosperms were still subordinate to gymnosperms and ferns except in coastal and riparian sites. In North America, angiosperms penetrated from the Gulf Coast to Alberta along the shores of Albian epicontinental seas and also first appeared in Alaska along shifting seashores. In river deposits, there was some segregation of fossils in different kinds of alluvium, ferns and gymnosperms being found in fine sediments and angiosperms being relatively abundant in coarser sediments deposited by high-energy streams. The leaves attributed to Platanus were commonly buried in sand.
Cenomanian
Patterns changed only slowly. Until late Cenomanian time, angiosperm pollen generally remained rare compared to gymnosperm pollen, especially in Gondwanaland. Retallack and Dilcher (1981b ) concluded that angiosperms were still mainly confined to coastal and riparian habitats, growing as mangroves along tidal distributaries, in swamps along lagoon margins, as scrub on natural levees, and as floating aquatics. Some of the richest Cenomanian fossil beds are in the Dakota Formation of Kansas, laid down in a variety of coastal and deltaic environments. Angiosperm leaf fossils there and elsewhere in North America and Europe show increasing diversity. Many have been questionably identified with living families.
Tricolpate dicot pollen continued to be widespread and appeared for the first time in high arctic latitudes. A derivative type, Normapolles , first appeared in the Cenomanian in eastern North America and Europe. It has been interpreted as signaling a reversion to wind pollination, perhaps by Fagaceae. Wind pollination implies large stands of a species, not just scattered seashore or streambank pioneers.
Turonian
Eastern North America and Europe were still in contact at high latitudes in Turonian time. They retained pollen floras that were broadly similar but already had distinct genera. In both regions Normapolles diversified rapidly. There, for the first time, angiosperm pollen was abundant relative to gymnosperm
pollen, suggesting extensive hardwood forests. The ancient, primitive Clavatipollenites had become rare except in Australia. Gondwanaland remained conifer dominated.
Coniacean—Campanian
Normapolles pollen, presumably from hardwood forests, continued to dominate eastern North America, Greenland, and Europe, which were still in contact during Coniacean—Campanian time. Western North America and eastern Asia had evidently come into contact; they shared a newly evolved pollen assemblage called Aquilapollenites . Also derived from the tricolpate type, Aquilapollenites was presumably dicot pollen, but it has no living counterpart. Some Aquilapollenites also appeared in Gondwanaland during this period. Another new derivative of the tricolpate dicot type, Proteacidites, appeared in Australia in the Coniacean; this may mark the appearance of the Proteaceae, another family in the Rosidae. The Proteaceae are a classic example of a group whose modern distribution suggests a Gondwanaland origin. In the Coniacean, it would have still been possible for the Proteaceae to migrate over land between Australia and Africa. By the Santonian, Africa was isolated.
The Gondwanaland migrations of the southern beech, Nothofagus, are especially intriguing (Axelrod and Raven 1982; Schuster 1976). A member of the Fagaceae and almost certainly originally from the northern hemisphere, Nothofagus appeared in the fossil record in South America, Australia, and New Zealand in Senonian time. Its progenitor had evidently migrated from North to South America during one of the intermittent Cretaceous contacts and had continued through Gondwanaland before the link between South America and Australia via Antarctica was broken. It is unlikely that Nothofagus could have made this migration except over a continuous land path. The seed dispersal radius of living Nothofagus is very short, and the trees are dependent on symbiotic mycorrhizal fungi that are not dispersed with the seed.
Maestrichtian
The sharp distinction between the angiosperm floras of eastern North America—Europe and western North America—eastern Asia lasted into the early Maestrichtian. The discreteness of these provinces was destroyed in the late Maestrichtian, when the epicontinental seas that had bisected the continents withdrew. The pollen record then shows general mixing of Normapolles and Aquilapollenites taxa.
From the Maestrichtian on, paleobotanists are less cautious about assigning
angiosperm fossils to living families and even genera. Muller (1981) recognized 31 extant families as present in the Maestrichtian, including those noted above as present in earlier periods.
It is customary to characterize Cretaceous angiosperm evolution as explosive, with some justification. Hughes (1976) estimated that at the start of the Cretaceous, the world had about 3,000 species of vascular plants, roughly half of them ferns and half gymnosperms. He estimated that by the Maestrichtian, there were about 2,500 surviving ferns and gymnosperms and about 20,000 species of angiosperms. The explosion had scarcely begun then; the world now has about 285,000 angiosperm species in about 300 families.
The diversification of the angiosperms is commonly credited to their specialized symbioses with animal pollinators and seed dispersers. Escape from wind pollination and wind dispersal allowed origin and survival of species growing in special scattered habitats and forming complex associations rather than dominating extensive, simple stands. What little evidence there is on Cretaceous angiosperm reproduction, however, suggests that angiosperms were then mostly generalists, with pollen dispersed by wind and unspecialized insects and seeds dispersed largely by wind and water. By the Maestrichtian, some coadaptation for pollination and dispersal by specific animal groups was evidently finally beginning, but major diversification of the most important modern pollinators and dispersers, namely butterflies, moths, bees, birds, and bats, was still far in the future.
Cretaceous—Tertiary Transition
(Hickey 1984; Pillmore et al. 1984; Smit and Kaars 1984; Tschudy and Tschudy 1986; Wolfe and Upchurch in press)
Controversy is currently very active over a hypothesis that the Cretaceous ended with a bang: a huge asteroid impact caused clouds of debris to darken the earth and shut down photosynthesis, resulting in mass mortality and extinctions of many marine and terrestrial species. There is no doubt that there were major changes in flora and fauna at the Cretaceous—Tertiary transition. The argument is over the causes and whether the changes were nearly instantaneous or spread out over perhaps 10,000 to 100,000 years or more.
So far, only a little evidence on both sides of the controversy has been adduced from the fossil record of seed plants.
Evidence from various sites in the Rocky Mountain region suggests that several fossil pollen taxa, including most kinds of Aquilapollenites, disappeared abruptly at the end of the Cretaceous. Disappearance of the angiosperm
pollen was followed by a temporary increase of fern spores from less than 30% to nearly 90% of the microfossil record. According to Wolfe and Upchurch (in press), the subsequent return to dominance of angiosperms followed a pattern of normal ecological succession but at a slower rate; this implies migration from survivors outside the region.
Axelrod (personal communication 1985) suggested that such changes might have been due to regional volcanic ashfalls rather than a global phenomenon. Hickey (1984) described floristic changes at the Cretaceous—Tertiary transition as geographically variable and gradual. Aquilapollenites taxa reached their peak diversity at the end of the Maestrichtian, but their decline extended through the Paleocene until extinction in the Eocene, a span of about 10 million years. In many other regions of the world, Hickey (1984) described floristic changes at the end of the Cretaceous as slight to moderate, with Cretaceous plant species persisting after dinosaur extinctions.
The argument is not over.
Paleogene (Paleocene, Eocene, And Oligocene
(Axelrod 1984; Axelrod and Raven 1978, 1985; Daghlian 1981; Dilcher 1973; Germeraad et al. 1968; Hickey 1984; Hos 1975; Kemp and Harris 1975; Lakhanpal 1970; Leopold 1969; Muller 1981; Penny 1969; Raven and Axelrod 1974; Stebbins 1981; Tanai 1972; Tiffney 1985; Tschudy 1984; Wolfe 1972)
Whatever extinctions took place at the Cretaceous—Tertiary transition, a variety of seed plant lineages came through and radiated rapidly in the Paleogene, when a great many living genera of conifers and angiosperms first appear in the fossil record.
The global configuration of continental contacts and separations changed drastically during the Paleogene. The North Atlantic continued to drive an oceanic wedge northward, and by the Oligocene, North America—Greenland and Europe were separated. Asia and Alaska, however, remained in contact, sea levels remained low, and there may have been no other major oceanic barriers to migration in Laurasia. In contrast, the breakup of Gondwanaland became complete during the Paleogene. Africa was an island through the whole period, except for possible brief contact with Eurasia. India separated from Madagascar early in the Paleocene and moved northward to collide with Asia in the Eocene. By late Eocene or early Oligocene, South America, Antarctica, and Australia had separated and Australia was moving into lower latitudes.
For coastal angiosperms, the oceans probably continued to be pathways
of migration rather than barriers. For example, Nypa, a mangrove swamp palm, which had first entered the fossil record in the East Indies in Late Cretaceous time, evidently migrated westward by the Tethys Sea route and across the widening Atlantic Ocean. In the Paleocene and Eocene, Nypa pollen and its sea-borne fruits were present in Southeast Asia, Australia, Europe, West Africa, and on the Atlantic coasts of both North and South America. Nypa is not known to have survived outside the tropical Indo-Pacific Ocean region after the Oligocene. Eocene pollen of Nypa and other mangroves, including Avicennia, Rhizophora, and Sonneratia, and of the beach tree, Barringtonia, has been found in southwestern Australia, which was situated south of 50°S at that time; this is a much higher latitude than is reached by any modern mangroves. Rhizophora was also present in the Paleogene of Borneo and the Caribbean. Likewise, there are widely scattered Eocene fossils attributed to various modern genera of pantropical beach plants, such as Caesalpinia in Nigeria and South America.
Transoceanic dispersal appears unlikely for the forest conifers and broadleaf hardwoods often assigned to the Arcto-Tertiary geoflora, whose ranges around the northern hemisphere were discussed previously. Some of these broad ranges, for example, of Sequoiadendron, Magnolia, and Platanus, may date from the Cretaceous, but many of the genera are not known from before the Paleogene. Perhaps some of them crossed between eastern North America and Europe in the early Paleogene before the Atlantic barrier developed, while others crossed by the longer available Bering land bridge. The relative importance of these two routes is still highly speculative. There is no evidence that the Arcto-Tertiary geoflora originated in a limited region and time span and spread as a unit. Presumably it was assembled gradually by convergence of taxa migrating over differing pathways from a variety of geographic origins. It may eventually be possible to sort out their migrational histories, but this awaits more and better dated fossil records.
There is an intriguing possibility that the northward drift of the Indian subcontinent ferried some seed plant lineages from Gondwanaland to Asia. Some Indian Paleocene and Eocene fossil floras are known from coastal and freshwater sediments associated with Deccan Traps lava flows. Some of these fossils are attributed to Podocarpus, a genus that survives in India today. Living Indian podocarps, however, belong to sections of the genus unknown in Africa or Madagascar and believed to have immigrated to southern Asia via the East Indies from Australia in much later Neogene time. Also, several of the Paleogene Indian fossils were attributed to genera of sea-dispersed coastal plants for which no ferry would have been needed, such as Nypa, Cocos, Barringtonia, Sonneratia, and Terminalia . Some of the Deccan fossils, however, belong to taxa not adapted for ocean dispersal, including Musaceae, which have a wide disjunction between Africa, Madagascar, and Southeast Asia.
Deep sea drilling has brought up Paleogene fossils from now sunken Indian Ocean regions: Broken Ridge, then perhaps part of Australia, and Ninetyeast Ridge, then perhaps a chain of volcanic islands. The floras included Araucaria, Casuarina, Proteaceae, and other taxa then present in Australia. Early Eocene floras in Australia show that Araucaria, Podocarpus, Casuarina, Nothofagus, and many genera of Proteaceae grew as far south as 60°S paleolatitude. Axelrod (1984) discussed the problem of explaining the presence of evergreen trees in polar regions. In the Oligocene, Eucalyptus and Leptospermum appeared in the Australian fossil record. It is uncertain whether their progenitors were present but undetected in the old Gondwana flora or whether they immigrated from the tropical Indo-Pacific islands.