Avian Use of Marshes on the Lower Colorado River[1]
Bertin W. Anderson, Robert D. Ohmart, Julie K. Meents, and William C. Hunter[2]
Abstract.—Vegetation composition and structure was used in designating eight types of marshes along the lower Colorado River. Marsh-types that had high densities of cattail (Typhalatifolia ) and bulrush (Scirpusacutus ), supported more wading birds (including Yuma Clapper Rail [Ralluslongirostris yumanensis ]) and insectivores. Waterbirds and shorebirds favored more open marsh situations. The interface between marsh and terrestrial riparian habitat was more important to terrestrial birds than to marsh birds.
Introduction
Marshes are an important component of wildlife habitat in riparian systems. In the lower Colorado River valley, marshes frequently occur adjacent to terrestrial and aquatic habitats and may support avian species characteristic of both habitats, such as the endangered Yuma Clapper Rail (Ralluslongirostrisyumanensis ), which occurs primarily in marsh vegetation.
In the lower Colorado River valley, marshes differ in vegetation composition and structure; these differences are reflected in the avian species which inhabit them. In this paper, we describe the different types of marshes and their dominant vegetation characteristics. Marsh use by granivores (GR), shorebirds (SB), insectivores (I), waterbirds (WB), and wading birds (WA) is evaluated by marsh-type and by season.
Methods
A total of 30 transects was established in representative marsh vegetation along the Colorado River between Davis Dam, Arizona/Nevada border and the United States/Mexico International Boundary (fig. 1). Each transect was censused three times per month between May 1976 and July 1978, using a modified variable transect method (Emlen 1971; Anderson and Ohmart 1977). Results of all censuses on a transect in a particular season were averaged to obtain seasonal densities. Seasons, designated to coincide with changes in avian population, included winter (December–February), spring (March–April), summer (May–July), late summer (August–September), and fall (October–November). Bird species found in marshes were combined into groups or guilds on the basis of similarity in feeding behavior and general habitat use (table 1).
Foliage height diversity (FHD) and foliage density were measured by the method of MacArthur and MacArthur (1961). Percent dominance of each plant species was evaluated by determining the number of times each species was included in foliage density measurements. Principal components analysis (PCA) (Nie etal .) was used to determine which vegetation variables were highly intercorrelated; the PCA also provided a series of new, noncorrelated variables based on the original data. Each transect was given a score representing its relative position on the continuum represented by each of these derived variables. These scores were used in grouping transects into general marsh-types.
Results and Discussion
Vegetation
The PCA indicated that several of the vegetation variables measured were highly correlated. Cat-tail (Typha latifolia ) and bulrush (Scirpusacutus ) tended to occur together, but neither coexisted with phragmites (Phragmitesaustralis ). These three variables were combined into a derived variable; the relative amounts of these plant species are represented by the score of each transect (Fig. 2). On the
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Bertin W. Anderson is Faculty Research Associate; Robert D. Ohmart is Associate Director; Julie K. Meents is Research Biologist; William C. Hunter is Field Biologist; all are at the Center for Environmental Studies, Arizona State University, Tempe, Arizona.
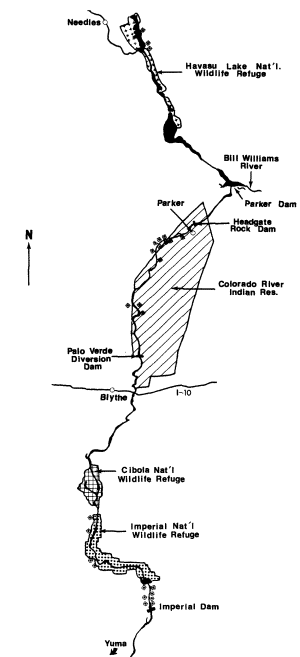
Figure l.
Map of marsh transects along the lower Colorado
River.

first principal component or derived variable, positive scores indicate increasing amounts of cat-tail or bulrush; negative scores indicate increasing amounts of phragmites. Structural differences, derived from FHD measurements, indicated that cat-tail/bulrush tends to be short (below 1.5-m. height) and dense, whereas phragmites is taller (above 1.5-m. height) and more spatially heterogeneous.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The PCA also showed that trees (mostly salt cedar [Tamarixchinensis ]) and grasses tend to occur together. A transect's score on this derived variable reflects its measured density of trees and grasses (fig. 3). A negative score indicates few or no trees and grasses and a positive score indicates above-average amounts of trees and grasses.
Marsh-Type
The relative scores of the transects on these two derived variable suggested that there were basically six types of marshes on the lower Colorado River. Two additional marsh-types, an open (water) marsh and a geographically isolated marsh, were compared with the original six types.
The types were:
Type I—nearly 100% cat-tail/bulrush, small amounts of phragmites and open water;
Type II—nearly 75% cat-tail/bulrush, many trees and grasses interspersed;
Type III—about 25–50% cat-tail/bulrush, some phragmites, open water, some trees and grass;
Type IV—about 35–50% cat-tail/bulrush, many trees and grasses interspersed;
Type V—about 50–75% cat-tail/bulrush, few trees and grasses interspersed;
Type VI—nearly 100% phragmites, little open water;
Type VII—open marsh (75% water), adjacent to sparse marsh vegetation; includes sandbars and mudflats when Colorado River is low; and
Type VIII—Topock Marsh near Needles, California; vegetatively similar to Type I, but with even denser stands of bulrushes. This marsh-type was kept separate for purposes of analysis as it was censused for only five months and is geographically isolated from the rest of the study area.
Avian Occurrence in Marshes
Granivores
Highest densities of granivores occurred in most marsh-types during fall, winter, and spring (table 2). Occurrence of granivores in marshes was related to the occurrence of trees and grasses. Marsh-type II, having high occurrences of both cat-tail and bulrush and trees and grasses, would be expected to supply the largest food source for granivores. This type supported the highest density of granivores. Type IV, with average densities of cat-tail/bulrush and above-average densities of trees and grasses, and Type III, with average cat-tail/bulrush and average trees and grasses, had the second and third highest granivore densities, respectively. The lowest densities of granivores occurred in Type VI (phragmites), which had few cat-tails, bulrushes, trees, or grasses, and thus provided limited food sources. However, in Type VI, highest densities of granivores occurred during summer. Upon reviewing the species composition for this season and type, the Brown-headed Cowbird (Molothru s ater ) was the sole species involved and
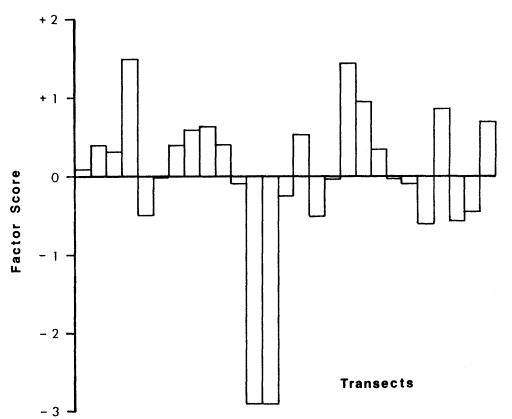
Figure 2.
Factor score of each transect on a derived variable representing the amount of cat-tail,
bulrush, and phragmites. Positive scores indicate increasing amounts of cat-tail and
bulrush; negative scores indicate increasing amounts of phragmites.
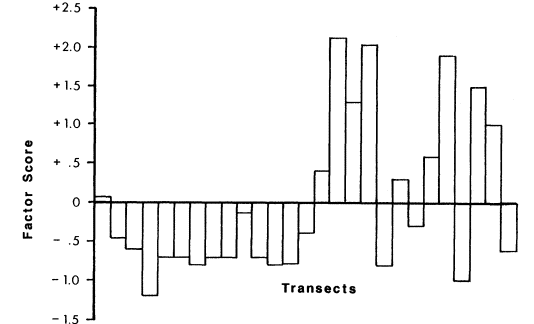
Figure 3.
Factor score of each transect on a derived variable representing density of trees and
amount of grass. Positive scores indicate above-average amounts of trees and
grasses; negative scores indicate few or no trees and grasses.
probably acted as a nest parasite rather than actually foraging on seeds.
Shorebirds
Shorebirds were most numerous during fall, winter, and spring (table 2). The highest densities occurred in open marsh (Type VII) where exposed sandbars and mudflats facilitated shorebird use. All other marsh-types had low densities; practically no shorebirds occurred in the most vegetationally dense types (I, II, V, and VI).
Insectivores
Insectivore analysis was complex due to the large number of species and the high niche separation of these species. However, the following trends were evident (table 2). Seasonally, the majority of marsh-types supported highest densities of insectivores in fall and winter. Dense and intermediately dense cat-tail/bulrush (Types I and V) did not follow this trend; highest densities of insectivores in these types occurred during summer.
Cat-tail/bulrush marshes of Types II and III had the highest insectivore densities at all seasons (especially fall and winter for Type II; see table 2). Type III had remarkably even numbers of insectivores at all seasons. These consisted mostly of Red-winged Blackbird, Song Sparrow, Common Yellowthroat, and Long-billed Marsh Wren during all seasons. Cat-tail/bulrush marshes of Type I and marshes with trees and grasses interspersed (Type V) had lower insectivore densities and followed Type III in being even at all seasons.
Though only censused during spring and summer, the very dense Type VIII marsh had very high total densities, consisting primarily of Longbilled Marsh Wren, Common Yellowthroat, and Song Sparrow. Overall for breeding insectivores, as cat-tail/bulrush became very dense, densities of Long-billed Marsh Wren, Common Yellowthroat, and Song Sparrow increased (noted in Types VIII and I). As cat-tail/bulrush approached average density, numbers of Red-winged Blackbird increased (note especially Type III; table 2). Overall, large numbers of Yellow-rumped Warbler contributed to the high fall, winter, and spring insectivore densities in most marsh-types. In open marsh (Type VI), however, high densities were accounted for by large numbers of the Water Pipit, which requires more open areas for foraging.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Waterbirds
Seasonally, waterbirds were most numerous during fall, winter, and spring in all marshtypes. Waterbirds reached their highest densities in open marsh (Type VII; table 2). Marsh-types with trees and grasses interspersed (V) and those consisting of phragmites (VI) had very low waterbird densities. The most obvious trend was for waterbirds to avoid densely vegetated areas.
However, Type VIII, a dense cat-tail/bulrush marsh, had unexpectedly high waterbird densities during spring. The American Coot was the overwhelmingly most numerous species. Though the apparent trend for denser vegetation to have
lower waterbird densities is probably realistic, it must be tempered with the realization that detectability of species in this guild is difficult due to visibility problems and the lack of highly vocal species (with the exception of the American Coot). More open vegetation would facilitate easier feeding for most species, while denser vegetation would provide cover when species in this guild were not feeding.
Wading Birds
Since the collective total for wading birds was much less than that for any other guild, alterations of marshlands could cause a severe decline in their numbers (table 2). Thus, wading birds may provide an accurate gauge for ascertaining the health of marshlands along the lower Colorado River. One species from this group, the Yuma Clapper Rail, will be analyzed in detail in the following section.
Seasonally, wading birds were most numerous in spring, summer, and late summer (table 2). Most wading birds moved out of marshland after breeding, as indicated by differences between breeding (spring and summer) and wintering seasons in most marsh-types. During summer, cattail/bulrush marshes of Types V, VIII, and I, and phragmites marshes of Type VI, in that order, were the most important types of vegetation for breeding. For wading birds collectively, the species comprising the vegetation do not appear to be as important as its density. Open marsh (Type VII) had the lowest density of wading birds. Although not taken into account in this paper, the presence or absence of tall, dead trees should be considered for any studies on habitat use for breeding herons and cormorants, as they use the dead trees for nesting platforms.
Yuma Clapper Rail
The Yuma Clapper Rail is endemic to the marshes of the lower Colorado River from Needles, California south to the Colorado Delta of Mexico, including immediate drainage areas (Salton Sea, Gila River, and Salt River). An analysis of marsh use of the Yuma Clapper Rail was necessary because of its close relationship with marshes and because of the threatened status of this subspecies. The analysis covered only spring, summer, and late summer, as the majority of the Colorado River population is migratory (Smith 1975).
Clapper Rail densities were compared for each of the defined marsh-types by month (table 3). The importance of the different marsh-types was determined by ranking. During each month, the marsh-type with the highest rail density received the highest score; the marsh-type with the lowest density received the lowest score. Scores for each marsh-type were then added across months and compared directly.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Type V was by far the marsh-type most highly used by the rails. Type I ranked one-half as high as Type V, but was nearly double the rank of Types II, III, VI, and VIII. Marsh-types with trees and grasses interspersed (Type IV) and very open marshes (Type VII) consistently ranked low. Of interest, Type VI (mostly phragmites) had relatively high ranking during spring when rails arrive to breed, and Type VII (above-average density of cat-tail/bulrush and above-average density of trees and grasses) had very high ranking during late summer when rails dispersed from the breeding areas. But during summer breeding, these marsh-types were not used extensively by rails. Even though rails appear to select certain marsh-types for actual breeding, nonbreeding marginal areas may still be important.
Conclusions
Among the six avian categories or guilds considered, five occurred in significantly greater densities in at least one marsh-type, relative to the density in the marsh-type with the lowest density. Thus, the first conclusion is that the species within these guilds have definite habitat preferences. Among the six guilds, peak populations were reached in four different marsh-types. Thus, the second conclusion is that to maximize densities and diversities of birds using marsh as habitat, it is necessasry to have a variety of marsh-types available.
A third conclusion is that among the marshtypes studied, dense stands of phragmites supported the lowest avian densities and diversities. But even this marsh-type was potentially valuable to the Yuma Clapper Rail in the prebreeding season and to granivorous species, especially in winter, spring, and late summer.
Unrestricted vehicular traffic has been found to reduce the use of marsh and open-water areas by waterfowl (Ohmart and Anderson 1978; Anderson and Ohmart 1981). One simple means of habitat improvement would be to close roads adjacent to at least some marshes during periods of avian peak use, i.e., winter.
While man has created a large portion of the existing marshes along the lower Colorado River (Ohmart etal . 1975), we hasten to point out that channelization and riprapping activities, in general, reduce the quality of marshes (ibid .). Riprapping and channelization activities should be undertaken only if the creation of marsh habitat is part of a mitigation proposal.
Immediate action must be taken to support the mesquite (Prosopisglandulosa ) skeletons in Topock Marsh to ensure a nesting area for the heron and cormorant rookery, which may well be the largest remaining rookery along the lower Colorado River.
Channelization projects usually result in destruction of sandbars and mudflats. Our final conclusion is that when these operations occur, an assertive stance must be taken to ensure that mitigation includes plans for creation of new sandbars and mudflats in the general areas.
Acknowledgments
We would like to thank Helen Wood for presenting this paper. Susan M. Cook and Jane R. Durham provided editorial assistance and Marcelett Ector typed the various drafts of the manuscript. Cindy D. Zisner typed the final draft. Judy Huff, Janet Jackson, Ann Kasprzyk, and Helen Wood constructed the tables and graphs. Rodney H. Ohmart and Elaine Hassinger prepared the final figures. We wish to give special acknowledgment to Alton Higgins who was instrumental in conducting the field work. This study was supported by USDI Bureau of Reclamation Contract Number 7-07-30-V0009.
Literature Cited
Anderson, B.W., and R.D. Ohmart. 1977. Climatological and physical characteristics affecting avian population estimates in Southwestern riparian communities. p. 193–200. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitats: a symposium. [Tucson, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Anderson, B.W., and R.D. Ohmart. 1981. Vegetation management. Final report to USDI Bureau of Reclamation, Boulder City, Nevada. In preparation.
Emlen, J.T. 1971. Population densities of birds derived from transect counts. Auk 88:323–342.
MacArthur, R.H., and J.W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.
Nie, N.H., J.G. Jenkins, K. Steinbrenner, D.H. Bent. 1975. Statistical package for the social sciences. McGraw-Hill, Inc., New York, N.Y. 675 p.
Ohmart, R.D., and B.W. Anderson. 1979. Wildlife use values of wetlands in the arid southwestern United States. p. 278–295. In : P.E. Greeson, J.R. Clark, and J.E. Clark (ed.). Wetland functions and values: the state of our understanding. Proceedings of the national symposium on wetlands. [Minneapolis, Minn., November 1978]. American Water Resources Association, Minneapolis, Minn. 674 p.
Ohmart, R.D., W.O. Deason, and S.J. Freeland. 1975. Dynamics of marshland formation and succession along the lower Colorado River and their importance and management problems as related to wildlife in the arid Southwest. p. 240–251. In : Transactions of the 40th North American Wildlife and Natural Resources Conference. Wildlife Management Institute, Washington, D.C.
Smith, P.M. 1975. Habitat requirements and observations on the Clapper Rail (Ralluslongirostrisyumanensis ). Unpublished M.S. Thesis, Arizona State University, Tempe, Ariz. 35 p.