Riparian Bird Community Structure and Dynamics
Dog Island, Red Bluff, California[1]
Stephen A. Laymon[2]
Abstract.—This 5-year weekly census on an 18-ha. plot documents the seasonal and year to year changes in a riparian bird community. High bird densities were found in all seasons. Species richness and bird density were found to be higher than in other wooded habitats in the West. The community was analysed using the concept of species diversity. Effects of Brown-headed Cowbird parasitism and the "habitat island" nature of the study area are discussed.
Introduction
"In California, the habitat that most clearly approximates the eastern broadleaved hardwood forests is the riparian woodland. This is so because of the nature of the trees in this woodland, their denseness, and the unparalleled diversity of the bird life." (Small 1974).
"Today, with the last extensive remnants of these forests in jeopardy, it behooves us to weigh the importance of riparian habitat to birds and other wildlife." (Gaines 1977).
These two quotations address both the importance of, and the threat to, lowland riparian systems in California and the West. Statewide, the extensive riparian forests encompassing hundreds of thousands of hectares have been reduced to mere remnants within 100 years.
The forests of the interior of California had not been explored extensively by ornithologists until recent years. The summer heat and winter fog were discouraging factors. One study in Red Bluff and the surrounding area retreated to the mountains in mid-May as the temperatures increased (Grinnell etal . 1930), thus missing many late migrant and nesting birds. It was 1964 before mention was made of the massive numbers of fall migrants using the Sacramento Valley willow thickets (Chase and DeBenedictis 1964). In 1974 when I began this study, the only quantitative evidence of riparian bird populations consisted of eight breeding bird censuses and one winter bird study (Dembosz et al . 1972; Gaines 1973; Ingles 1950; Manolis 1973; Shuford 1973; Tangren 1971; Tangren 1972; Winkler 1973a, 1973b). Nothing quantitative had been published on migration.
Since 1974 much more information on riparian bird populations has appeared. In 1979 and 1980 alone, American Birds (vol. 33 and 34) published seven breeding bird censuses and 10 winter bird population studies from riparian areas in California. Several short-term government agency studies in the Sacramento Valley have resulted in administrative reports and papers of varying quality (Stone 1976;[3] Brumley 1979; Michny etal . 1975; Hehnke and Stone 1978; Hurst etal . 1980). Most of these studies suffered due to lack of trained field ornithologists to gather the field data. The most impressive study has been conducted along the lower Colorado River, coordinated by Ohmart and Anderson (e.g. Anderson and Ohmart 1977). Their study area represents a different avifauna than that found in the Central Valley.
The present study was formulated to gather population data on a regular basis, monitor changes in breeding and winter populations from year to year, and document the extent of migratory bird usage of the lowland riparian woodland vegetation.
Study Area
Dog Island City Park is located on the west side of the Sacramento River at the north end of the city of Red Bluff, in central Tehama County, California. Development of the park has been held to a low level with parking lot, restrooms, a few picnic tables, and trails. The area is
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Stephen A. Laymon is a Graduate Student in the Department of Forestry and Resource Management, University of California, Berkeley.
[3] Stone, T.B. 1976. Birds in riparian habitat of the upper Sacramento River. The Resources Agency, Department of Fish and Game memorandum report. Sacramento, Calif.
17.7 ha. in extent and consists of a 5.9 ha. island, 9.4 ha. mainland portion and 2.4 ha. of river channel (see fig. 1).
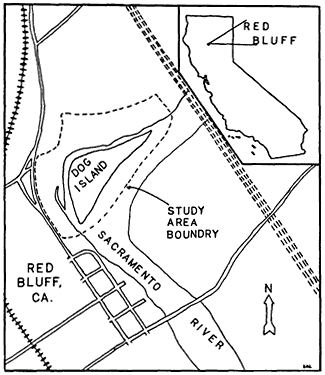
Figure l.
Dog Island study area and environs.
The climate of the study area is moderate Mediterranean, with a wet, cool winter and a long, dry, hot summer. During wet winters, much of the area is inundated, sometimes for several weeks at a time.
The dominant vegetation-type is floodplain riparian woodland. The vegetation is quite diverse with different species being dominant on portions of the area, often forming clumps or bands of a single species.
Vegetation was characterized using a series of 2 m.-wide belt transects through the system. The average canopy closure, determined from the belt transects, was 80% while the average groundcover was 68%. The mean canopy height was 13.4 m. (range 2–35 m.) and total density was 1,619 trees per ha. The most frequent trees were willow (Salix , seven spp., 300 per ha.), black walnut (Juglans sp., 296 per ha.), box elder (Acernegundo , 288 per ha.), Oregon ash (Fraxinuslatifolia , 154 per ha.), white alder (Alnusrhombifolia , 124 per ha.), and Fremont cottonwood (Populusfremontii , 110 per ha.). Percent canopy/understory by species was walnut 21%, cottonwood 20%, box elder 18%, willow 16%, alder 8%, and ash 7%. The groundcover consisted of mugwort (Artemesiadouglasiana ), blackberry (Rubus , three spp.) and various grasses. There was a small marsh in the center of the island.
The area is a vegetational island surrounded by blue oak (Quercusdouglasii ) woodland, urban development, grassland, and orchards. The nearest extensive riparian woodland is 15 km. to the south. Additional information on the area is presented in Laymon (1983).
Methods
Sampling Methods
The census route was laid out along existing pathways which made loops through the two main portions of the study area. The census route was 3.9 km. in length and gave good coverage of the area. No attempt was made to estimate actual numbers except for breeding pairs, using a supplementary spot-mapping system (Kendeigh 1944) and an extrapolation of migrant populations for selected species.
Censuses were begun between 06:30 and 08:30. Censuses ran an average of 170.5 minutes (range 115 to 270 minutes). Census time averaged longer in spring and fall when migrants were present. Censuses were run weekly except when the area was flooded. When possible, days of high wind or rain were avoided. The census method consisted of walking slowly along the route and stopping at regular spots. All birds both seen and heard were recorded. Birds flying overhead were recorded but a notation to that effect was made.
A total of 238 censuses were conducted between February 1974 and June 1980. Of these only the 197 censuses taken from 1975 through 1979 are used in this presentation.
Data Analysis
A system of numbered weeks, starting 1–7 January was used to organize the data. All censuses taken during each week were pooled to gain the overall trend. Week 2 (8–14 Jan.) was not represented by any censuses so the figures for that week are the average of weeks 1 and 3. A three-number running average was used as a curve-smoothing device on the composite graph.
The total number of individuals (TI) and the frequency of occurrence (F) was calculated for all species seen from 1976 through 1979. The species were then ranked by a composite figure computed by multiplying TI by F (proportion of 1.0).
Species diversity (H') was calculated using the Shannon formula (Shannon and Weaver 1948). The maximum species diversity (H'max) was also calculated for each sample as was evenness (J'= H'/H'max) (Pielou 1966). Foliage height diversity (FHD) was calculated using the same formula based on presence or absence of foliage at three levels (0 to 2 m., 2 to 5 m., 5+ m.) at 700 points.
The nesting pair densities were based on spot-mapping of singing males and located nests.
Density was figured for a hypothetical km2 of similar habitat. A similar method was used for winter populations but the numbers were based on an average of all censuses from December through mid-February. The migration numbers for selected species were obtained from individual-year census data. The total number of migrants was estimated based on a three-to-four day layover for building up fat stores during migration.4
Results
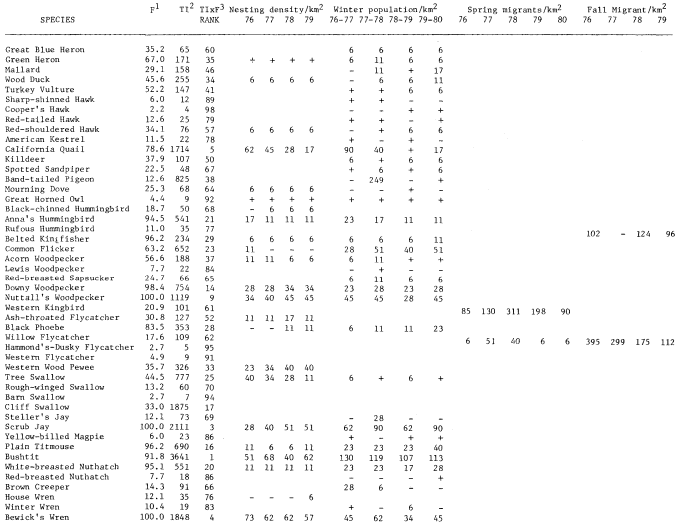
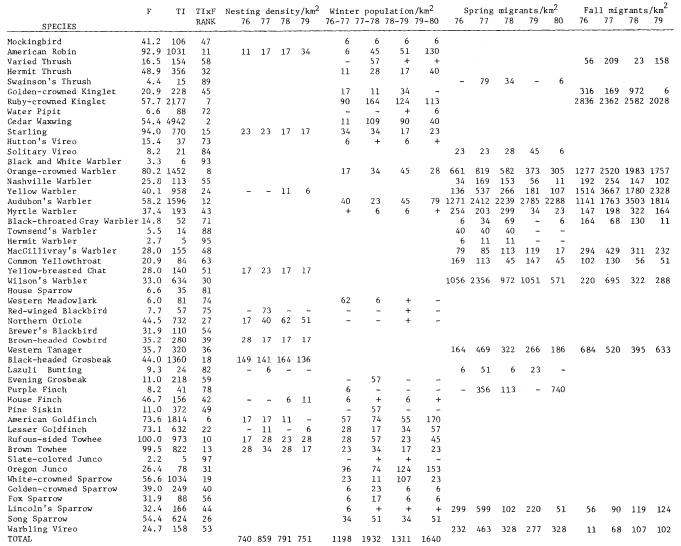
A total of 147 species of birds were recorded during the censuses. The 98 species listed in table 1 occurred regularly. This table shows TI, F, Rank, and nesting, winter, and estimated migrant populations. The two forms of Yellow-rumped Warbler and Dark-eyed Junco are treated as separate taxa. Scientific names of the birds are not included to save space.
Species Diversity
Species diversity (H') is a concept which indicates both the richness and evenness of the members of a plant or animal community. It is mainly used to compare the diversity of one system or area to another.
Robert MacArthur (1972) used this formula to compare bird species diversity (BSD) with foliage height diversity (FHD) (MacArthur and MacArthur 1961). He found that FHD explained BSD better than plant species diversity or any other habitat parameter that he measured. This theory has been widely tested and has held in many cases (Balda 1975).
|
|
Species diversity of the nesting pairs on the study area was figured for each year, 1976 through 1979 (table 2). The average for the four years was 3.00 and the FHD was 1.08. All nesting BSD figures were higher than predicted using MacArthur's graph. This is not at all surprising since the study area was 17.7 ha., as compared to 2-ha. areas used by MacArthur (1972). A larger area will generally have more species nesting, and therefore will have a larger BSD. A sample of three 2-ha. plots was chosen from the 1979 nesting census data. BSD figures of 2.05, 2.40, and 2.70 were calculated for those plots. The latter two figures fit into MacArthur's linear regression, but the first figure falls well below any of his results. It is possible that the FHD was not the same for these small plots as for the whole study area.
Comparison of BSD at different seasons is also possible. When comparing the nesting and wintering population BSD, the wintering BSD was considerably higher, but shows less evenness. This is misleading due to the averaging of 6–12 census dates to obtain the wintering population figures. In order to make valid comparisons between winter and nesting populations, I chose two winter and two summer census weeks and again obtained a higher BSD on the winter samples. This was due mainly to a higher number of species, since the evenness was very similar (table 2).
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
BSD's for spring and fall census weeks were also calculated. A great deal of difference from week to week for both BSD and evenness was obtained. Overall, summer had the lowest BSD and spring, the highest. This is not surprising since the spring census weeks had the highest number of species and the summer census weeks had the lowest (table 2). When comparing weeks 16 (16–22 April) and 19 (7–13 May), the BSD was much lower during week 19 even though the species total was the same due to the large influx of Cedar Waxwings. Increased numbers of one species will always decrease the BSD.
Seasonal Changes
Figure 2 shows the changes in numbers of birds and in the total and average numbers of species from week to week, averaged over the combined 5-year period of study. The total number of species is a summation of all species found during each week for the five years of the study. The averge number of species is the average number recorded on all censuses during each week over the 5-year period. The peaks and dips in the three lines do not occur in the same places.
Total number of species (species) and average number of individuals (individuals) held relativey constant from the beginning of the year through week 5 (29 Jan.–4 Feb.). In week 6 (5–11 Feb.) and 7 (12–18 Feb.) individuals dropped while species held steady. The drop in individuals was due to a reduction in Band-tailed Pigeons in 1978. During week 8 (19–25 Feb.) individuals began to climb, reaching a peak on week 10 (5–11 Mar.). This peak was due to large increases in swallows, Audubon's Warblers, and American Goldfinches.
Species began to swing upward on week 9 (26 Feb.–4 Mar.) and rose to a peak on weeks 15 to 17 (9–29 Apr.) as spring migration was in full swing. Species dropped from weeks 17 (23–29 Apr.) through 24 (11–17 June) as the wintering species left and migrants passed through, leaving only the summer residents.
On the other hand, individuals dropped from week 10 (5–11 Mar.) through week 14 (2–8 Apr.) as the numerous Band-tailed Pigeons, Audubon's Warblers, and American Goldfinches left the area. Individuals then rose again through week 18 (30 Apr.–6 May) with migrant Cedar Waxwings, Wilson's Warblers, and Black-headed Grosbeaks making large increases. Individuals declined steadily through week 24 (11–17 June) as the migrants and winter residents left.
On week 26 (25 June–1 July) both individuals and species began to rise due to the fledging of young and influx of the first fall migrants. On week 28 (9–15 July), with species still rising, individuals began to fall as the swallows left. Individual continued to fall through week 33 (13–19 Aug.) as the common summer residents (Western Wood Pewee, Northern Oriole, and Black-headed Grosbeak) left. At the same time, species continued to rise as early migrants moved through.
Species peaked from weeks 35 to 37 (27 Aug.–16 Sept.) and then again on week 43 (22–28 Oct.) as the early and late waves of migrants passed
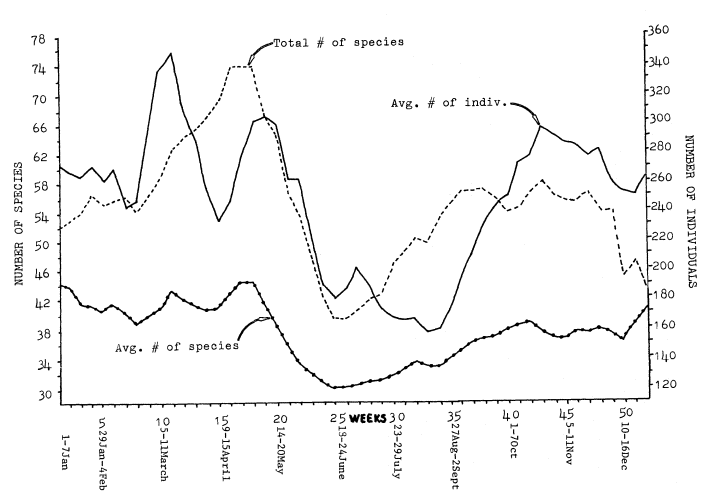
Figure 2.
Variations in numbers of species and individuals (three-week running average) for 1975 through 1979.
through. Individuals began to rise on week 34 (20–26 Aug.) and continued to go up until a peak on week 43 (22–28 Oct.) was reached. This was caused first by large numbers of Orange-crowned Warbler and Yellow Warbler, and later by an in-flux of Ruby-crowned Kinglet, Cedar Waxwing, and Audubon's Warbler. Individuals and species then dropped gradually to week 50 (10–16 Dec.).
The difference between the average number of species and total number of species was greatest during migration, and smallest during winter and summer. This was caused by a greater chance of rarities during migration, and differences in migration timing from year to year. An average of not more that 45 species was found on any week, but the chances of new species occurring on censuses during that week in succeeding years were greatest during migration.
Nesting Season
The lowland riparian nesting avifauna in central California is quite rich. A total of 83 species have been documented as nesting in the riparian areas of the Sacramento Valley in the past 100 years. Five species (Common Merganser, Spotted Sandpiper, Western Wood Pewee, Starling, and Brown-headed Cowbird) have colonized the area; and 11 species (Double-crested Cormorant, Cooper's Hawk, Bald Eagle, Long-eared Owl, Willow Flycatcher, Western Flycatcher, Purple Martin, Swainson's Thrush, Blue-gray Gnatcatcher, Bell's Vireo) have become either very rare or extirpated during that time (Gaines 1974).
Of these 83 species, 35 were found breeding on Dog Island during the study interval. An average of 29.5 species nested each year (range 27 to 31). Twenty-five species nested in all four years, two nested in three years, four nested in two years, and four nested once. Of the 10 species not found every year, five (Black-
chinned Hummingbird, Black Phoebe, House Wren, Yellow Warbler, and House Finch) colonized the area during the study. Two species (Common Flicker and American Goldfinch) became extinct as nesting species, and three species (Red-winged Blackbird, Lazuli Bunting, and Lesser Goldfinch) both colonized and became extinct as nesting species during the study.
The most abundant nesting species were the Black-headed Grosbeak, Bewick's Wren, and Bushtit. The total number of nesting birds averaged 139 pairs in 1977. In most cases, the numbers of nesting pairs of each species varied little from year to year. Notable exceptions were California Quail and Tree Swallow which declined; and Western Wood Pewee, Scrub Jay, American Robin, and Northern Oriole, which increased.
The origin of the nesting pairs was 50% residents, 13% short-distance migrants, and 37% long-distance migrants. The origin of the 35 nesting species was 44% residents, 28% short-distance migrants, and 28% long-distance migrants. Of the unstable species (i.e., those colonizing or becoming extinct), four were long-distance migrants. Only the Black Phoebe was a resident. This indicates a very unstable situation for migrants, with 45% colonizing or becoming extinct in four years.
Nesting guilds were devised on the basis of the type of nesting situation used. The avian community nesting guild composition is presented in table 3. The proportion nesting on the ground was quite low, considering the dense groundcover. This was not surprising however, since ground nesting is dangerous (Skutch 1976). The proportion of tree-hole nesters was high. This was undoubtedly a result of the large number of snags and decadent trees on the plot. These provided sites for woodpeckers to excavate their homes which in turn provided homes for the secondary cavity nesters. Many of the live trees also had natural cavities from broken-off limbs.
| |||||||||||||||||||||
Spring and Fall Migration
Each year gave a different overall impression of migration. Much of this impression came from the presence or absence of irruptive species, but much also came from the actual timing and numbers of migrants.
To gain some insight into the spring migration differences over the years, I used two criteria: 1) migration timing; and 2) actual numbers. The species accounts (Laymon 1981) and estimated spring migrant numbers (table 1) were examined for six categories: 1) record high totals; 2) record low totals; 3) record early arrival date; 4) record late arrival date; 5) record early departure date; and 6) record late departure date. The number of species that set records in any category in any year was recorded (table 4). From a birdwatcher's standpoint, 1977 was clearly the "best", since more species arrived earlier, left later, and set records for numbers. Just as clearly, 1980 was the "worst" spring, with more species arriving later, leaving earlier, and setting record low numbers. The three years 1976 through 1979 all had many early and three late departure dates, indicating a mixed situation.
Fall migration was analysed in the same manner (table 4). Late and early arrival and departure dates seemed to have little relation to the general timing of migration in fall. This was because fall migration is a much more relaxed affair. Migration timing is much less precise than in the spring. As a result, very little pattern was revealed in this portion of the analysis except that 1979 had the most early arrivals and the fewest late arrivals, and 1976 was just the reverse. The record high and low numbers revealed that 1977 had the most highs and no lows, 1976 and 1979 had the least highs and the most lows.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Winter Populations
Winter populations varied from one year to the next, much more so than breeding populations. The difference between the highest and lowest breeding years was only 14%, while the difference between the highest and lowest wintering populations was 38%. The higher totals in 1977–78 were caused almost exclusively by irruptive species such as Band-tailed Pigeon, Steller's Jay, Varied Thrush, Cedar Waxwing, Evening Grosbeak, and Pine Siskin.
Examination of a sample of 25 regularly occurring, non-irruptive species revealed that the winter of 1979–80 actually had the highest wintering population, with an average of 229 individuals per survey. The winter of 1976–77 had 160 individuals, 1977–78 had 198 and 1978–79 had 178 individuals per survey. This was still a 30% difference between the highest and lowest years. Variations in food supply, temperature, rainfall, and floodwaters must have been responsible for some of the variation in bird numbers. Much may also have been caused by nesting success and food supply on the breeding grounds.
Discussion
Yearly Changes in Numbers and Migration Timing
Numbers of migrants, both individuals and species, varied greatly from year to year. The cause of these changes was not readily evident. In some cases, the immediate weather patterns, such as rain or wind, probably grounded many migrants or kept them from leaving an area. In other cases, long-range weather patterns such as a drought, late snows, or a long, severe cold spell probably changed migration patterns. In 1977, many more than average numbers of migrants were found at Dog Island. This was a drought year in which little winter rain and no spring rain fell. It was likely that the spring migrants avoided the foothills which were parched by mid-March, and depended heavily on the lowland riparian forest for a refueling stop. It was also likely that the heavy fall migration that year was caused by the mountain springs and meadows being dry by late summer. The abundant food in the lowland riparian zone seems to be especially important to migrants during drought years.
Another factor leading to changes in numbers of migrants from year to year is nesting and wintering success. The number of birds passing north in the spring is the number which survived the winter. Winter, not breeding season, has been suggested as the season which limits bird population numbers in the temperate zone (Fretwell 1972). It is possible that destruction of riparian woodland to the south and the removal of vast areas of tropical forest may limit nesting and migrant populations here in the future. Removal of riparian woodland here may now be limiting nesting populations to the north. The birds migrating south in the fall are the sum of the surviving adults which went north to breed plus their offspring. In years of good nesting success, the number of migrants could be much higher that in poor years.
Effects of Brown-headed Cowbirds
Brown-headed Cowbirds are a recent immigrant to the Sacramento Valley of California. This bird is a parasitic species which lays its eggs in other birds' nests. The host species then incubates the eggs and raises the young, leaving the cowbird free to look for additional nests and food. The arrival of the cowbird has had a drastic effect on the native avifauna throughout California, particularly in lowland riparian systems adjacent to agricultural areas (Gaines 1974).
Had this study been done prior to the arrival of the Brown-headed Cowbird, a remarkably different avian community would probably have been present. Certain species such as the Willow Flycatcher, Western Flycatcher, Swainson's Thrush, Blue-gray Gnatcatcher, Bell's Vireo, and Warbling Vireo nested in the lowland riparian areas. None of these are now known to nest anywhere in lowland interior California. Other species such as the Yellow-billed Cuckoo, Yellow Warbler, Common Yellowthroat, and Song Sparrow still nest in the Sacramento Valley, but are now found in reduced numbers. With the exception of the Yellow-billed Cuckoo, Western Flycatcher, and Swainson's Thrush, all of the species in both groups are considered to have high to very high susceptibility to cowbird parasitism (Gaines 1974). Evidence implicating the Brown-headed Cowbird in the decline of these species is only circumstantial, since no studies were conducted during the 1940's and 1950's as the decline was occurring. However, no other theory presently explains the decline.
Of all the species which have declined or were extirpated, only the Yellow Warbler nested in the study area during the study. I believe that numbers of nesting individuals, numbers of nesting species, and species diversity were much lower in the study area due to cowbird parasitism. The species which have maintained high populations are either hole-nesters, species which reject cowbird eggs (Rothstein 1971), or species such as the Black-headed Grosbeak, which are large enough to raise a cowbird young along with their own. The small open-cup nesting species which accept cowbird eggs have all declined or disappeared completely.
Effects of the Habitat Island Nature of the Study Area
The study area is an island of riparian vegetation in a sea of farmland, oak woodland, and a river. The theories of island biogeography were developed by studying oceanic islands. These systems have higher extinction rates, lower species diversity, and fewer species than a comparable area of mainland (Diamond 1969). Distinction has been made between oceanic and habitat islands (MacArthur 1972). Habitat islands do not have as marked an effect on avian community structure, but the effect is still evident (Vuilleumier 1970).
Extinctions and colonizations of 10 nesting bird species were recorded in the four summers of the study. This represented almost a third of the nesting species, and seems like a high rate of turnover. BSD figures were at or below the predicted values using MacArthur's FHD linear
regression. Both of these findings may be related to the habitat island situation.
As more and more habitat islands are created by conversion of riparian forest to other uses, this study area will become more representative of the valley forests. If the islands are large enough, the effect will not be disastrous since the migrant riparian species in the West have been accustomed to travelling over hundreds of kilometers of desert to nest along the streams. Resident species may not fare as well, and none will fare well if the islands are too small.
An additional topic to consider regarding habitat islands is the concentration of migrants on these islands. This concentration has been documented in desert oases and on coastal islands such as the Farallons on the central California coast. It is very likely that the study area drew a much greater number of migrants because it was surrounded by miles of inhospitable habitat for forest birds than it would if a similar 18-ha. plot was found within a much more extensive stand of forest.
Values of Riparian Systems
The value of a vegetation-type can be measured in a number of ways. Some of the most common methods are bird densities, species richness, and the number of species unique to the system. These figures can be compared from system to system and a relative importance can be derived. Using American Birds breeding bird census and winter bird population studies for 1979 and 1980 I compiled table 5 which compares Dog Island results with other riparian areas and other vegetation-types in the West. No figures for migrant densities are available in the literature, so no comparisons were made.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Breeding bird densities were 24% lower at Dog Island than at 14 other riparian sites, while the number of species was slightly higher. When compared to other vegetation-types, however, the Dog Island figures were much higher, with bird densities ranging from 47% higher than chaparral to 89% higher that desert scrub. All other vegetation-types also had fewer species, ranging from 33% less in chaparral to 80% less in desert scrub. The Dog Island winter density figures are comparable to other riparian sites (5% less) and range from 57% higher than oak woodland to 89% higher than desert scrub. Winter bird species numbers were 58% higher than other riparian sites, 67% highr than oak woodland, and 85% higher that desert scrub. None of these other vegetation-types even begin to approach the riparian sites in either density or species richness.
The third category, number of species unique to the vegetation-type, is a harder one to compare. Little information has been gathered on densities of the same species in different vegetation-types, so habitat preferences are usually a matter of subjective judgment. Twenty of the regularly occurring species: Wood Duck, Common Merganser, Red-shouldered Hawk, Spotted Sandpiper, Downy Woodpecker, Black Phoebe, Willow Flycatcher, Tree Swallow, House Wren, Swainson's Thrush, Ruby-crowned Kinglet, Warbling Vireo, Yellow Warbler, Yellow-breasted Chat, Wilson's Warbler, Lazuli Bunting, Black-headed Grosbeak, American Goldfinch, Lincoln's Sparrow, and Song Sparrow are believed to have their highest densities in riparian systems. Of this group, Wood Duck, Red-shouldered Hawk, Willow Flycatcher, Swainson's Thrush, Yellow Warbler, Yellow-breasted Chat, Wilson's Warbler, and American Goldfinch are rarely found away from this vegetation-type. Is is doubtful that other vegetation-types in the West could match these figures.
In this study we have seen that a great many individuals of a great many species of birds use the Dog Island riparian system. This 17.7-ha. plot receives a great deal of avian use throughout the year. It is vital to the nesting, migrating, and wintering success of numerous individuals of many species. When viewed on a larger scale, the riparian systems of the West are vital to the maintenance of many populations, subspecies (Blue Grosbeak, Red-shouldered Hawk, and Yellow-billed Cuckoo), and, in a few cases, species of birds (Willow Flycatcher and Bell's Vireo). These systems have a high species diversity throughout the year. They are an essential link for long-distance migrants from the north and are an important wintering ground for many species.
Literature Cited
Anderson, B.W., and R.D. Ohmart. 1977. Vegetation structure and bird use in the lower Colorado Valley. p. 23–24. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: a symposium. [Tuscon, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43. 217 p. Rocky Mountain Forest and Range Experiment Station, Fort Collins Colo.
Balda, R.P. 1975. Vegetation structure and breeding bird diversity. p. 59–80. In : D.R. Smith (tech. coord.). Management of forest and range habitats for nongame birds: Proceedings of the symposium. [Tuscon, Ariz., May 6–9, 1975]. USDA Forest Service GTR-WO-1. 343 p. USDA Forest Service, Washington, D.C.
Brumley, T.D. 1976. Upper Butte Basin study 1974–75. Wildlife Management Branch administrative report 76–1. The Resources Agency, Department of Fish and Game, Sacramento, Calif.
Chase, T. Jr, and P. DeBenedictis. 1964. Middle Pacific Coast region report. Audubon Field Notes 19:71.
Dembosz, D., K. Ficket, and T. Manolis. 1972. Disturbed floodplain woodland. Amer. Birds 26:978–979.
Diamond, J.M. 1969. Avifaunal equilibria and species turnover rates on the Channel Islands of California. Nat. Acad. Sci., Proc. 64:57–63.
Fretwell, S.D. 1972. Populations in a seasonal environment. Princeton University Press, Princeton, N.J.
Gaines, D. 1973. Floodplain riparian woodland. Amer. Birds 27:995.
Gaines, D.A. 1974. A new look at the nesting riparian avifauna of the Sacramento Valley, California. Western Birds 5:61–80.
Gaines, D.A. 1977. The valley riparian forests of California: their importance to bird populations. p. 57–85. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Grinnel, J., J. Dixon, and J.M. Linsdale. 1930. Vertebrate natural history of a section of Northern California through the Lassen Peak region. University of California Publ. in Zool. 35(1).
Hehnke, M., and C.P. Stone. 1978. Values of riparian vegetation to avian populations along the Sacramento River system. p. 228–235. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Ga., December 11–13, 1978]. USDI Forest Service GTR-WO-12. 410 p. Washington, D.C.
Hurst, E., M. Hehnke, and C.C. Goude. 1980. The destruction of riparian vegetation and its impact on the avian wildlife in the Sacramento River Valley, California. Amer. Birds 34:8–12.
Ingles, L. 1950. Nesting birds of the willow-cottonwood community in California. Auk 67:325–332.
Kendeigh, S.C. 1944. Measurement of bird populations. Ecol. Monogr. 14:67–106.
Laymon, S.A. 1981. Avifauna of an island of lowland riparian woodland: Dog Island City Park, Red Bluff, California. Unpublished masters thesis, California State University, Chico.
Laymon, Stephen A. 1983. Photodocumentation of vegetation and landform change on a riparian site, 1880–1980: Dog Island, Red Bluff, California. In : R.E. Warner and K.M. Hendrix (ed.). California Riparian Systems. [University of California, Davis, September 17–19, 1981]. University of California Press, Berkeley.
MacArthur, R.H. 1972. Geographical ecology. Harper and Row, Inc. New York, New York.
MacArthur, R.H.; and J.W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.
Manolis, T. 1973. Disturbed floodplain woodland. Amer. Birds 27:994–995.
Michny, F.J., D. Boos, and F. Wernette. 1975. Riparian habitats and avian densities along the Sacramento River. The Resources Agency, Department of Fish and Game, Wildlife Management Branch administrative report 75–1.
Pielou, E.C. 1966. Species diversity and pattern diversity in the study of ecological succession. J. Theoret. Biol. 10:370–383.
Root, R.B. 1967. The niche exploitation pattern of the Blue-gray Gnatcatcher. Ecol. Monogr. 37:317–350.
Rothstein, S.I. 1971. Observation and experiment in the analysis of interactions between brood parasites and their hosts. Amer. Natur. 105:71–74.
Shannon, C.E., and W. Weaver. 1948. The mathematical theory of communication. University of Illinois Press. Urbana, Ill.
Shuford, D. 1973. Disturbed riparian stream border. Amer. Birds 27:1005.
Skutch, A.F. 1976. Parent birds and their young. University of Texas Press. Austin, Texas.
Small, A. 1974. The birds of California. Winchester Press. New York, New York.
Tangren, G. 1971. Riparian oak woodland. Amer. Birds 25:967–969.
Tangren, G. 1972. Riparian oak woodland. Amer. Birds 26:977–978.
Vuilleumier, F. 1970. Insular biogeography in continental regions. The northern Andes of South America. Amer. Nat. 104:373–388.
Winkler, D. 1973a. Riparian oak woodland. Amer. Birds 27:689.
Winkler, D. 1973b. Riparian oak woodland. Amer. Birds 27:996.