13—
RIPARIAN SYSTEMS AND THE ECOLOGY OF BIRD POPULATIONS

Seasonal Variation of Bird Numbers in a Riparian Forest, Sacramento Valley, California[1]
Robert S. Motroni[2]
Abstract.—During the period 1 September 1976 to 30 August 1977, breeding and migratory birds were censused bi-weekly in a riparian system located 21 km. south of Yuba City, Sutter County, California. A significantly greater number of birds utilized the plant associations of the 174-ha. study area during the fall and winter (September to February) as opposed to the spring and summer (March to August). Forage substrates and food sources present on a seasonal basis were utilized by migratory species. The importance of early seral stage riparian growth to total bird numbers was significant. Habitat management in riparian systems of the Sacramento Valley should be directed toward wintering populations in addition to the requirements of breeding riparian bird species.
Introduction
Riparian systems are frequently considered one of the most productive forms of wildlife habitat in North America. Many bird species are wholly, or at least partially, dependent on riparian plant communities to perpetuate their kind. Johnson et al . (1977) found that of 166 breeding species in west Texas, southern Arizona, and southern New Mexico, 77% were partially dependent and 51% completely dependent upon riparian plant communities. Similar figures are available for California, where 43% of the species breeding in riparian zones dominated by cottonwood and willow exhibit a "primary affinity" to this vegetation-type (Gaines 1977). Cottonwood stands along the Verde River of Arizona provide the only breeding habitat for over 50% of the breeding species found there (Carothers and Johnson 1975).
Riparian systems are equally important to winter migrants and residents. Until recently however, the importance of wintering habitat has been neglected by researchers even though this season may be the critical one for bird populations. Anderson and Ohmart (1977, 1979) suggested that the mean niche breadth of all bird species along the lower Colorado River was narrowest with respect to use of vegetative structure in winter and was accompanied by reduced habitat overlap. This indicates that winter may be the period of greatest stress to bird populations in this area.
Riparian systems are clearly one of the most important types of wildlife habitat in North America. However, most of the significant literature on them has appeared only during the last 10 years. Of the 254 papers included in a recent bibliography on the importance of riparian systems to terrestrial wildlife, 85% were from the decade of the 1970's (Motroni 1980). The present paper deals with the seasonal variation of bird numbers within plant communities of a riparian system based on seral stage, total numbers, species composition, and foraging guild.
Study Area and Methods
Study Area
The riparian system selected for study was the 174-ha. Bobelaine Audubon Sanctuary, located on the west bank of the Feather River directly opposite the confluence of the Feather and Bear Rivers. This riparian system is approximately 21 km. south of Yuba City, Sutter County, California (38° 35' 45" west longitude). The study area, located in a region dominated by agriculture, is bordered on the west by a slough that is the byproduct of a borrow pit created in the early 1900's during levee construction. The eastern edge of the study area abuts the Feather River for a distance of 3.25 km.
Five major plant associations were found on the study area and represent various stages of plant succession. Plant associations were iden-
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Robert S. Motroni is Wildlife Biologist, USDA Forest Service, Plumas National Forest, Blairsden, Calif.
tified based on the most abundant and dominant plant species present and are described below.
1. Riparian Shrub 1 (RS1)—composed primarily of California rose (Rosacalifornica ), Mexican tea (Chenopodiumambrosioides ), willow (Salix spp.), and blackberry (Rubusvitifolius ).
2. Riparian Shrub 2 (RS2)—dominated by willow (Salix spp.), button bush (Cephalanthusoccidentalis ), and various species of annual and perennial grasses.
3. Cottonwood Forest (CF)—contains cottonwood (Populusfremontii ), and valley oak (Quercuslobata ) with an understory of Mexican tea, poison oak (Rhus diversiloba ) and various grasses.
4. Riparian Forest (RF)—dominated, in contrast to the Cottonwood Forest association, by a dense stand of cottonwood with an understory of poison oak, blackberry, wild grape (Vitiscalifornica ), blue vervain (Verbenahastata ) and other herbs and grasses.
5. Oak Woodland (OW)—on areas of high ground dominated by mature valley oak, sycamore (Platanusracemosa ), and cottonwood. The understory is composed of widely separated shrubs such as poison oak and California rose, interspersed with grass. This association has an open, park-like appearance. A more complete quantitative description of the plant associations examined can be found in Motroni (1979).
Methods
Plot census techniques were utilized after the establishment of study plots in areas representative of the various plant associations of the study area. Plots of varying size depending on vegetation limits were established in each of the five plant associations: RS1—5 ha.; RS2—2.5 ha.; CF—3 ha.; RF—2 ha.; and OW—3 ha.
Plot censuses were conducted on alternate weeks from the initiation of the study on 1 September 1976 to its completion on 30 August 1977. Two methods of data collection and interpretation were utilized during the study. During the wintering and migratory periods, when little if any territoriality was evident in the avian population, a modified form of the summation method (originally a breeding bird census technique) was used. This method has been reported by Palmgren (1930) and Udvardy (1947), and was evaluated by Enemar (1959). The modified summation method entails recording the number and species of all birds encountered over the entire census plot. When the data were interpreted, an estimation of abundance and bird habitat preference on the day of the census was obtained.
During the breeding season, after territory establishment, the spot-map method described by Kendeigh (1944), and Enemar (1959), was utilized but modified such that all birds encountered, not only to territorial males or females, were marked on a plot map during each visit. Data collected in this manner were interpreted by compiling sightings of each species onto a separate species map. This facilitated an analysis of territory size, habitat preference, and recogninition of non-territorial transient birds. Also, as Williamson (1964) has pointed out, the mapping results provided information on the distribution and numbers of various species as well as on their interrelationships with the entire avian community. These spot-map censuses were initiated at the first sign of territorial activity and were made at regular intervals throughout the breeding season in order to cover the peak song periods of all species encountered. Data were collected only on fair days due to the adverse effects of wind and rain on the song and activity patterns of birds.
Bird species diversity was calculated with the Shannon-Wiener Diversity Index (Cox 1972) utilizing logs to the base e.
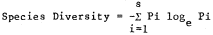
where Pi equals the decimal fraction of total individuals belonging to the ith species and s equals the total number of species. Maximum diversity (H maximum), an expression of the highest diversity value possible assuming equal numbers of all species, and evenness (E) were also obtained from the diversity formula. The seasonal distribution of the total number of birds observed was tested with the Kruskal-Wallis one-way analysis of variance by ranks described by Siegel (1956).
Bird species encountered during the fall/winter and spring/summer census periods were placed in foraging guilds based primarily upon the type of substrate and major food type utilized. A guild has been defined by Root (1967) as: "a group of species that exploit the same class of environmental resources in a similar way." The foraging guild taxonomy to which species were assigned was patterned after Bent (1923–1958), Gaines (1977), and field observations. Those species that belonged to two foraging guilds (e.g. Townsends Warbler (Dendroicatownsendi ) and Rufous-sided Towhee (Pipiloerythrophalmus )) were treated in the same manner as Gaines (ibid .), in that both guilds were weighted equally. All species were classified within the following foraging guilds: omnivore; ground seed; ground insect; ground mammal; bark insect; foliage seed; foliage insect; and air insect.
Results
Plot Census of Avifauna
Fall/Winter
Fall/winter bird censusing was conducted from 12 September 1976 to 25 February 1977 on a
bi-weekly basis, for a total of 12 census visits. Fifty-seven species of birds were observed.
The average density of birds ranged from 1,553 birds per 40 ha. in RS1 to 468 birds per 40 ha. in RS2. The high value obtained for RS1 was due primarily to the large numbers of Whitecrowned Sparrows (Zonotrichialeucophrys ) and Golden-crowned Sparrows (Zonotrichiaatricapilla ) utilizing this plant association during the wintering period. Bird species diversity for the fall/winter period of all plots sampled varied from a high of 2.99 in the RF to a low of 1.36 in RS1 (table 1).
| ||||||||||||||||||||||||||||||||||||||||||
Spring/Summer
Breeding bird censuses were conducted from 11 March 1977 to 30 August 1977 on a bi-weekly basis for a total of 13 census trips. Sixty-two species of birds were observed during this time period. Twenty-six of these species were common enough that sufficient data could be collected to make a breeding bird density determination.
Non-breeding birds observed on the plots, and those not belonging to any established territory, were classified as surplus birds in the plot sample data. The total number of breeding birds per 40 ha. ranged from a high of 1,000 in RF to a low of 368 in RS1 (table 2). Breeding bird species diversity was calculated for this period in the same manner as the wintering bird species diversity. However, since wintering bird numbers were calculated from direct observations, and breeding bird numbers (aside from birds classified as transient) were based on numerous observations of territorial individuals, a reevaluation of breeding bird data was necessary so that the diversities from these two time periods could be compared. This reevaluation was accomplished by counting the total number of observations of birds exhibiting territorial behavior, including females and transients, noted on the composite plot map for each species. Thus, diversity figures were based on 257 birds observed in RF; 344 in CF; 447 in OW; 398 in RS1; and 133 in RS2. Although females, due to their secretive nature in this particular time period, were not encountered as frequently as during the wintering period, the increased conspicuousness of active singing males was assumed to compensate in part for the lack of observation of females.
Bird species diversity values for the breeding period ranged from a high of 3.23 in OW to a low of 2.72 in CF (table 2). Evenness values ranged from a high of 0.833 in RF to a low of 0.760 in CF.
| ||||||||||||||||||||||||||||||||||||||||||||||||
Total Numbers
The total numbers of wintering versus breeding birds actually encountered on the various census plots are depicted in table 3. The vast majority of birds were observed during the
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
fall/winter census period (p < 0.05) with values ranging from 67% in the OW plot to 85% in the RS1 plot.
Foraging Guilds
The percentages of total breeding and wintering individuals in relation to their foraging guilds are depicted in table 4. Calculations of the total number of individuals in each foraging guild were done in the same manner as the diversity calculations for comparisons of breeding and wintering birds. This facilitated a more accurate comparison between fall/winter and spring/summer periods. Only those species known to forage in the plots sampled were included in the calculations.
During the fall/winter period, the majority of birds in each plant association fed primarily on the ground on seeds produced by shrubs and herbaceous vegetation, as well as on quiescent insect life. Bark insect guild birds were primarily permanent residents; the percentages of total numbers encountered during the census period were as follows: RF—75%; OW—77%; CF—66%; RS1—91%; and RS2—67%.
Conversely, migratory species (primarily wintering sparrows and the Dark-eyed Junco (Juncohyemalis ), dominated the ground seed guild, with the exception of the RF birds: RF—19%; OW—81%; CF—80%; RS1—84%; and RS2—56%. The high percentage of resident birds in the ground seed foraging guild in RF (81%) was due in part to habitat selection. The high densities of permanent resident birds such as California Quail (Lophortyx californicus ), Rufous-sided Towhee, and Brown Towhee (Pipilofuscus ) in this association overshadowed the presence of the migratory avifauna.
Resident species were again dominant in the ground insect foraging guild: RF—93%; OW—100%; CF—94%; RS1—99%; and RS2—100%. Similarly, species within the omnivore guild were composed entirely of resident birds in every plant association.
During the breeding period the utilization of insect life as forage increased markedly. Species found in the air insect guild were predominately migrants. The migratory composition of the foliage insect and air insect foraging guilds were, respectively, as follows: RF—71%, 100%; OW—43%, 96%; CF—27%, 100%; RS1—27%, 100%; and RS2—65%, 100%. The relatively low percentage of migratory birds in the foliage insect foraging guild was due in part to the presence of such residents as Bewicks Wren (Thryomanesbewickii ), Common Bushtit (Psaltriparus minimus ), and others that are flexible enough to change foraging guilds from the wintering to the breeding period.
If these species are excluded from foraging guild calculations then the percentages of migratory birds in the foliage insect guild ranged from 50–100% in the various associations surveyed. Gaines (1977) has shown that the foliage insect foraging guild is approximately equally divided between residents (53%) and migrants (47%) in a cottonwood/willow riparian forest. During the breeding period the bark insect guild continued to be dominated by resident birds: RF—98%; OW—89%; CF—100%; RS1—100%; and RS2—95%. As during the wintering period, the omnivore foraging guild was composed entirely of resident species in each plant association.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Discussion
Density and Seral Stage
The relationships between breeding bird populations and various aspects of plant succession have been examined by Johnston and Odum (1956), and Bond (1957). Diversity values and the numbers of birds supported in any particular plant association during the breeding season have been related to the seral stage of the plant association examined, and to the structural complexity of the vegetation. Breeding bird densities have also been related to the distribution and volume of tree foliage (Balda 1969, 1970). It is noteworthy that OW supported a lower
breeding bird density than those areas with relatively pure cottonwood stands such as RF. In contrast, those areas of CF that had a major valley oak component exhibited density trends comparable to OW (table 2).
The difference between breeding bird populations of the relatively pure cottonwood stands of RF and stands of OW may have been due to the more mesic conditions of the former and subsequent promotion of greater plant growth and higher invertebrate populations than were found in OW or CF. These mesic conditions, if present, also contributed to a more complex, stratified foliage profile. Gaines (1974) arrived at similar conclusions. Carothers etal . (1974), in a study of breeding riparian woodland bird populations in north-central Arizona, determined that those areas of relatively pure cottonwood stands supported the largest breeding bird populations, whereas areas of a mixed deciduous tree and shrub component supported smaller populations of breeding birds.
Breeding bird diversity values in those plant associations surveyed on the study area were consistently higher than the values for wintering birds (tables 1 and 2). This difference was a consequence of the large influx of a relatively few migrant species during the fall/winter period versus the presence of a more evenly distributed population of breeding species.
In contrast to the breeding period census results, the wintering density of OW was similar to that of RF (table 1). This similarity was due in part to the presence of migratory species in the bark insect and ground seed foraging guilds. The relatively open, xeric conditions of OW may have been more conducive to the growth of grasses and other seed-producing plants utilized by wintering species.
The large number of birds utilizing RS1 during the fall and winter was primarily due to seed-eating winter residents such as the Whitecrowned Sparrow and Golden-crowned Sparrow. This somewhat specialized habitat use by winter visitors has been previously described by Anderson and Ohmart (1977, 1980) for birds in the lower Colorado River Valley. In their study, winter visitors, and to a lesser degree summer visitors, exhibited an increase in specialization with regard to the use of vegetation-types. In addition, Fretwell (1972) predicted that winter residents will exhibit larger populations and a greater degree of habitat specialization than resident species, thus emphasizing the importance of this early seral stage association to wintering birds.
Total Numbers of Birds
It has been generally recognized that there are more birds utilizing the lowlands of California in fall and winter than in spring and summer. There is, however, little documentation of this dichotomy of seasonal utilization. Michny etal . (1975), in censusing an area of riparian woodland north of Princeton, Colusa County, California, discovered that during the month of August there was a mean of 23.8 birds per ha. Similar results were noted for May and June, with 24.0 birds per ha. However, this same area, when censused in February and March, produced a mean of 40.1 birds per ha. Winter visitor populations were observed to be about twice as dense as breeding bird populations in a south central Louisiana mature hardwood bottomland (Dickson 1978). Monthly winter populations varied between 1400 and 2000 birds per km.[2]
In contrast, other researchers have noted a higher level of avian use in riparian systems during the breeding period, for example, Anderson and Ohmart (1976) in several communities on the lower Colorado River; and Gavin and Sowls (1975) for a mesquite bosque (fide Szaro 1980). Factors influencing this disparity in use may include census technique, location of study area in relation to migratory pathways, habitat structure and condition, and suitability of adjacent areas.
During this study, a mean of 26.8 birds per ha. was found over the five associations surveyed during the fall/winter period, versus 7.7 birds per ha. during the spring/summer period. This three-fold increase in bird numbers during the fall and winter period may be due to a combination of several factors. During the wintering period there is a breakdown of territoriality among many menbers of the avian community. This facilitates an increase in gregariousness among many species and a consequent increase in number of birds inhabiting an area during the nonbreeding period. The presence of young-of-theyear in resident species also contributes to the size of the early fall population.
Nevertheless, the most important contribution to the fall/winter population size comes from the arrival of such abundant migrants as the American Robin (Turdusmigratorius ), Yellowrumped Warbler (Dendroicacoronata ), Rubycrowned Kinglet (Reguluscalendula ), Darkeyed Junco, White-crowned Sparrow, and Goldencrowned Sparrow. The general lack of strongly defended territories as well as the presence of an unpredictable and rigorous environment are considered to be primarily responsible for lower evenness levels exhibited by bird populations in winter than in other seasons (Tramer 1969; Holmes and Sturges 1975).
A number of explanations of the seasonality of bird habitat use are discernible and are of importance to habitat management. Species separated by ecological or geographic factors on wintering and breeding grounds merge during migration and contribute to higher densities along migration routes. Abramsky and Safriel (1980) suggested that the resultant increases in interindividual contacts and mortality during migration increase the importance of transition habitat. They concluded that migratory birds do not merely commute seasonally between two destinations, but rather occupy a series of transient
home ranges where food resources are heavily utilized.
Habitat structure and/or quality also provide a highly significant influence on seasonality of avian use. Anderson and Ohmart (1977) found that bird populations in riparian zones along the Colorado River reacted less in summer than during other seasons to such vegetation structure components as patchiness, foliage height diversity, etc. The importance of habitat quality inherent in riparian systems as it relates to adjacent system types and utilization by migratory birds has been frequently cited. Stevens etal . (1977) considered the importance of riparian systems to migrant birds in central Arizona to be substantial in that riparian systems contained up to 10.6 times the number of migrants per hectare as adjacent non-riparian areas. Similarly, Wauer (1977) suggested that riparian systems of the Rio Grande provide a major migratory route due primarily to the arid country through which these species pass.
The availability of winter food has been suggested by Fretwell (1972) to have marked influence on breeding densities via the maintenance of winter population levels. Species feeding primarily on seeds would, on the average, exhibit higher winter densities due to a more abundant food resource. The large number of wintering birds using seeds as a food source can be expected to occur primarily in the early seral stages of a plant community where such a food resource predominates. This was indeed the case in RS1 and the large number of seed eaters it supported during the wintering period. In addition, species of small body size might also be able to maintain high winter densities (Fretwell ibid .). The abundance of fringillids in this study during the fall/winter period supports the above theory.
Foraging Guilds
A general relationship between substrate, foraging guild, and migratory status can be discerned. It appears that winter residents and summer residents utilize those resources that are available only on a seasonal basis. During the wintering period ground seeds are in abundance and are used to a great extent by migratory species. Resident species dominate the ground insect and bark insect guilds, where the substrate and resource can be utilized on a yearround basis.
Conversely, during the spring/summer period, insectivores replace the granivores of the fall/winter period. The presence of foliage and active insect life provide a seasonal substrate and/or forage resource exploited by a separate group of migratory birds that includes such species as the Western Kingbird (Tyrannus verticalis ), Ash-throated Flycatcher (Myiarchus cinerascens ), Tree Swallow (Iridoprocnebicolor ), Northern Oriole (Icterusgalbula ), and Brown-headed Cowbird (Molothrusater ).
Similar results by Gaines (1977) showed that those substrates present on a year-round basis were used primarily by resident species, whereas most migratory species utilized seasonal substrates or food resources. Herrera (1978) examined an area dominated by extensive evergreen oak woodlands and concluded that resident species normally depend on the more permanent and less seasonal resources, in contrast to non-resident species whose subsistence is based on seasonally variable food supplies. Karr (1976a) described similar conclusions for a tropical bird community. To summarize, non-resident birds generally exploit an abundant, unpredictable food supply during both breeding and wintering periods (Willis 1966; Leck 1972; Bibby et al . 1976; Karr 1976b).
Summary and Conclusions
1. A significantly greater number of birds utilized five plant associations of the study area during the fall/winter period than in the spring/summer season. Winter resident species reached their greatest densities in early seral stage riparian growth.
2. Those forage strata and food sources present on a seasonal basis during the wintering and breeding periods were primarily utilized by migratory species.
The importance of early successional stage riparian vegetation to total bird numbers and overall species diversity on a seasonal level is significant. Recognition of this concept can influence managers to alter potential productivity of riparian systems during the planning and development of riparian reestablishment projects, levee maintenance, and bank protection projects. A number of the latter are capable of protecting birds while at the same time allowing natural riparian vegetation succession on river berms (indirect bank protection or river training devices as described by Keown etal . 1977).
To the casual observer, the breeding habitat of a species appears to be of prime importance. This is due in part to the disproportionate amount of effort by field workers in collecting data for this period. The three-fold increase in bird numbers utilizing riparian plant communities during the wintering and migratory periods as described earlier in this paper supports the concept that this ecosystem is of at least equal importance to the avifauna as a wintering area and migratory corridor.
Acknowledgments
Research was supported by a stipend from the Sacramento Chapter of the National Audubon Society and California State University, Sacramento. Thanks also go to Drs. Gary and Joan Fellers and Dr. M.D.F. Udvardy for their very helpful suggestions and critical reading of the manuscript. Research was conducted as partial
fulfillment of a graduate degree at California State University, Sacramento. Pegi Kelly and Diane Waterhouse typed the final manuscript.
Literature Cited
Abramsky, Z., and U. Safriel. 1980. Seasonal patterns in a Mediterranean bird community composed of transient, wintering and resident passerines. Ornis Scand. 11:201–216.
Anderson, B.W., and R.D. Ohmart. 1976. Wildlife use and densities report of birds and mammals in the lower Colorado River Valley. USDI Bureau of Reclamation, Lower Colorado River Region. 278 p. Boulder City, Nevada.
Anderson, B.W., and R.D. Ohmart. 1977. Vegetation structure and bird use in the lower Colorado River Valley. p. 23–34. In : R.R. Johnson and D.A. Jones (ed.). Importance, preservation and management of riparian habitat: A symposium. [Tucson, Arizona, July 9, 1977]. USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Anderson, B.W., and R.D. Ohmart. 1979. Riparian vegetation: an approach to mitigating for a disappearing habitat in the Southwest. p. 481–487. In : G.A. Swanson (tech. coord.). The mitigation symposium: A national workshop on mitigating losses of fish and wildlife habitat. USDA Forest Service GTR-RM-65, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 695 p.
Anderson, B.W., and R.D. Ohmart. 1980. Designing and developing a predictive model and testing a revegetated riparian community for southwestern birds. p. 434–450. In : R.M. DeGraff (tech. coord.). Management of western forests and grasslands for non-game birds. USDA Forest Service GTR-INT-86, Intermountain Forest and Range Experiment Station, Ogden, Utah. 535 p.
Balda, R.P. 1969. Foliage use by birds of the oak-juniper woodland and ponderosa pine forest in southeastern Arizona. Condor 71:399–412.
Balda, R.P. 1970. Effects of spring leaf-fall on composition and density of breeding birds in two southern Arizona woodlands. Condor 72:325–331.
Bent, A.C. 1923. Life histories of North American wildfowl. US National Museum Bulletin 126. Washington, D.C.
Bent, A.C. 1932. Life histories of North American gallinaceous birds. US National Museum Bulletin 162. Washington, D.C.
Bent, A.C. 1937. Life histories of North American birds of prey. US National Museum Bulletin 167. Washington, D.C.
Bent, A.C. 1939. Life histories of North American woodpeckers. US National Museum Bulletin 174. Washington, D.C.
Bent, A.C. 1940. Life histories of North American cuckoos, goatsuckers, hummingbirds and their allies. US National Museum Bulletin 176. Washington, D.C.
Bent, A.C. 1942. Life histories of North American flycatchers, swallows, larks, and their allies. US National Museum Bulletin 179. Washington, D.C.
Bent, A.C. 1948. Life histories of North American nuthatches, wrens, thrashers, and their allies. US National Museum Bulletin 195. Washington, D.C.
Bent, A.C. 1949. Life histories of North American thrushes, kinglets, and their allies. US National Museum Bulletin 196. Washington, D.C.
Bent, A.C. 1953. Life histories of North American wood warblers. US National Museum Bulletin 203. Washington, D.C.
Bent, A.C. 1958. Life histories of North American blackbirds, orioles, tanagers, and their allies. US National Museum Bulletin 211. Washington, D.C.
Bibby, C.J., R.E. Green, G. Pepler, and P.A. Pepler. 1976. Sedge Warbler migration and reed aphids. British Birds 69:384–399.
Bond, R.R. 1957. Ecological distribution of breeding birds in the upland forests of southern Wisconsin. Ecol. Monog. 27:351–382.
Carothers, S.W., and R.R. Johnson. 1975. Water management practices and their effects on nongame birds in range habitats. p. 210–222. In : D.R. Smith (tech. coord.). Proceedings of the symposium on management of forest and range habitats for nongame birds. USDA Forest Service GTR-WO-1, Washington, D.C.
Carothers, S.W., R.R. Johnson, and S.W. Aitchison. 1974. Population structure and social organization of southwestern riparian birds. Am. Zool. 14:97–108.
Cox, G.W. 1972. Laboratory manual of general ecology. 195 p. Wm. C. Brown Co., Dubuque, Iowa.
Dickson, J.G. 1978. Forest bird communities of the bottomland hardwoods. p. 66–73. In : R.M. DeGraff (tech. coord.). Proceedings of the workshop on management of southern forests for nongame birds. USDA Forest Service GTR-SE-14. 175 p. Southeast Forest Experiment Station, Ashville, North Carolina.
Enemar, A. 1959. On the determination of the size and composition of a passerine bird population during the breeding season. Var Fagelvarld, Suppl. 2:1–114.
Fretwell, S.D. 1972. Populations in a seasonal environment. 217 p. Princeton University Press, Princeton, N.J.
Gaines, D. 1974. A new look at the nesting riparian avifauna of the Sacramento Valley, California. Western Birds 5:61–80.
Gaines, D. 1977. The valley riparian forests of California: their importance to bird populations. p. 57–85. In : A. Sands (ed.). Riparian forests in California: their ecology and conservation. 122 p. Institute of Ecology, University of California, Davis.
Gavin, T.A., and L.K. Sowls. 1975. Avian fauna of a San Pedro valley mesquite forest. J. Ariz. Acad. Sci. 10:33–41.
Herrera, C.M. 1978. Ecological correlates of residence and non-residence in a Mediterranean passerine bird community. Jour. Anim. Ecol. 47:871–890.
Holmes, R.T., and F.W Sturges. 1975. Bird community dynamics and energetics in a northern hardwoods ecosystem. Jour. Anim. Ecol. 45:175–200.
Johnson, R.R., L.T. Haight, and J.M. Simpson. 1977. Endangered species vs. endangered habitats: a concept. p. 68–79. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: A symposium. [Tucson, Arizona, July 9, 1977]. USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Johnston, D.W., and E.P. Odum. 1956. Breeding bird populations in relation to plant succession in the Piedmont of Georgia. Ecology 37:50–62.
Karr, J.R. 1976a. Seasonality, resource availability, and community diversity in tropical bird communities. Amer. Nat. 100:973–994.
Karr, J.R. 1976b. On the relative abundance of migrants from the north temperate zone in tropical habitats. Wilson Bull. 88:443–458.
Kendeigh, S.C. 1944. Measurement of bird populations. Ecol Monog. 14:67–106.
Keown, M.P., N.R. Oswalt, E.B. Perry, and E.A. Dardeau, Jr. 1977. Literature survey and preliminary evaluation of streambank protection methods. US Army Corps of Engineers TR-H-77-9, US Army Corps of Engineers Waterways Experiment Station, Vicksburg, Miss. 262 p.
Leck, C.F. 1972. The impact of some North American migrants at fruiting trees in Panama. Auk 89:842–850.
Michny, F.J., D. Boos, and F. Wernette. 1975. Riparian habitats and avian densities along the Sacramento River. California Department of Fish and Game, Wildlife Management Branch Administrative Report 75–1. 42 p.
Motroni, R.S. 1979. Avian density and composition of a riparian forest, Sacramento Valley, California. M.S. Thesis, California State University, Sacramento. 172 p.
Motroni, R.S. 1980. The importance of riparian zones to terrestrial wildlife: an annotated bibliography. USDI Fish and Wildlife Service. Division of Ecological Services. 83 p. Sacramento, California.
Palmgren, P. 1930. Quantitative Untersuchungen uber die Vogelfauna in den Waldern Sudfinnlands. Acta Zool. Fennica 7:1–218.
Root, R.B. 1967. The niche exploitation pattern of the Blue-gray Gnatcatcher. Ecol. Monog. 37:317–350.
Siegel, S. 1956. Nonparametric statistics for the behavioral sciences. McGraw-Hill, New York, N.Y.
Stevens, L.E., B.T. Brown, J.M. Simpson, and R.R. Johnson. 1977. The importance of riparian habitat to migrating birds. p. 156–164. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: A symposium. [Tucson, Arizona, July 9, 1977]. USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Szaro, R.C. 1980. Factors influencing bird populations in southwestern riparian forests. p. 403–418. In : R.M. DeGraff (tech. coord.). Management of western forests and grasslands for non-game birds. USDA Forest Service GTR-INT-86, Intermountain Forest and Range Experiment Station, Ogden, Utah. 535 p.
Tramer, E.J. 1969. Bird species diversity: components of Shannon's formula. Ecology 50:927–929.
Udvardy, M.D.F. 1947. Methods of bird sociological survey, on the basis of some Tihany communities investigated. Arch. Biol. Hung. 17:61–89.
Wauer, H.R. 1977. Significance of Rio Grande riparian systems upon the avifauna. p. 165–174. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: A symposium. [Tuscon, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Williamson, K. 1964. Bird census work in woodland. Bird Study 11:1–22.
Willis, E.O. 1966. The role of migrant birds at swarms of army ants. Living Bird 5:187–231.
Riparian Bird Community Structure and Dynamics
Dog Island, Red Bluff, California[1]
Stephen A. Laymon[2]
Abstract.—This 5-year weekly census on an 18-ha. plot documents the seasonal and year to year changes in a riparian bird community. High bird densities were found in all seasons. Species richness and bird density were found to be higher than in other wooded habitats in the West. The community was analysed using the concept of species diversity. Effects of Brown-headed Cowbird parasitism and the "habitat island" nature of the study area are discussed.
Introduction
"In California, the habitat that most clearly approximates the eastern broadleaved hardwood forests is the riparian woodland. This is so because of the nature of the trees in this woodland, their denseness, and the unparalleled diversity of the bird life." (Small 1974).
"Today, with the last extensive remnants of these forests in jeopardy, it behooves us to weigh the importance of riparian habitat to birds and other wildlife." (Gaines 1977).
These two quotations address both the importance of, and the threat to, lowland riparian systems in California and the West. Statewide, the extensive riparian forests encompassing hundreds of thousands of hectares have been reduced to mere remnants within 100 years.
The forests of the interior of California had not been explored extensively by ornithologists until recent years. The summer heat and winter fog were discouraging factors. One study in Red Bluff and the surrounding area retreated to the mountains in mid-May as the temperatures increased (Grinnell etal . 1930), thus missing many late migrant and nesting birds. It was 1964 before mention was made of the massive numbers of fall migrants using the Sacramento Valley willow thickets (Chase and DeBenedictis 1964). In 1974 when I began this study, the only quantitative evidence of riparian bird populations consisted of eight breeding bird censuses and one winter bird study (Dembosz et al . 1972; Gaines 1973; Ingles 1950; Manolis 1973; Shuford 1973; Tangren 1971; Tangren 1972; Winkler 1973a, 1973b). Nothing quantitative had been published on migration.
Since 1974 much more information on riparian bird populations has appeared. In 1979 and 1980 alone, American Birds (vol. 33 and 34) published seven breeding bird censuses and 10 winter bird population studies from riparian areas in California. Several short-term government agency studies in the Sacramento Valley have resulted in administrative reports and papers of varying quality (Stone 1976;[3] Brumley 1979; Michny etal . 1975; Hehnke and Stone 1978; Hurst etal . 1980). Most of these studies suffered due to lack of trained field ornithologists to gather the field data. The most impressive study has been conducted along the lower Colorado River, coordinated by Ohmart and Anderson (e.g. Anderson and Ohmart 1977). Their study area represents a different avifauna than that found in the Central Valley.
The present study was formulated to gather population data on a regular basis, monitor changes in breeding and winter populations from year to year, and document the extent of migratory bird usage of the lowland riparian woodland vegetation.
Study Area
Dog Island City Park is located on the west side of the Sacramento River at the north end of the city of Red Bluff, in central Tehama County, California. Development of the park has been held to a low level with parking lot, restrooms, a few picnic tables, and trails. The area is
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Stephen A. Laymon is a Graduate Student in the Department of Forestry and Resource Management, University of California, Berkeley.
[3] Stone, T.B. 1976. Birds in riparian habitat of the upper Sacramento River. The Resources Agency, Department of Fish and Game memorandum report. Sacramento, Calif.
17.7 ha. in extent and consists of a 5.9 ha. island, 9.4 ha. mainland portion and 2.4 ha. of river channel (see fig. 1).
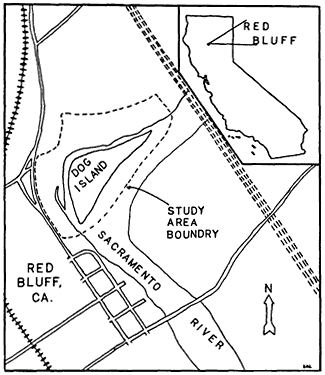
Figure l.
Dog Island study area and environs.
The climate of the study area is moderate Mediterranean, with a wet, cool winter and a long, dry, hot summer. During wet winters, much of the area is inundated, sometimes for several weeks at a time.
The dominant vegetation-type is floodplain riparian woodland. The vegetation is quite diverse with different species being dominant on portions of the area, often forming clumps or bands of a single species.
Vegetation was characterized using a series of 2 m.-wide belt transects through the system. The average canopy closure, determined from the belt transects, was 80% while the average groundcover was 68%. The mean canopy height was 13.4 m. (range 2–35 m.) and total density was 1,619 trees per ha. The most frequent trees were willow (Salix , seven spp., 300 per ha.), black walnut (Juglans sp., 296 per ha.), box elder (Acernegundo , 288 per ha.), Oregon ash (Fraxinuslatifolia , 154 per ha.), white alder (Alnusrhombifolia , 124 per ha.), and Fremont cottonwood (Populusfremontii , 110 per ha.). Percent canopy/understory by species was walnut 21%, cottonwood 20%, box elder 18%, willow 16%, alder 8%, and ash 7%. The groundcover consisted of mugwort (Artemesiadouglasiana ), blackberry (Rubus , three spp.) and various grasses. There was a small marsh in the center of the island.
The area is a vegetational island surrounded by blue oak (Quercusdouglasii ) woodland, urban development, grassland, and orchards. The nearest extensive riparian woodland is 15 km. to the south. Additional information on the area is presented in Laymon (1983).
Methods
Sampling Methods
The census route was laid out along existing pathways which made loops through the two main portions of the study area. The census route was 3.9 km. in length and gave good coverage of the area. No attempt was made to estimate actual numbers except for breeding pairs, using a supplementary spot-mapping system (Kendeigh 1944) and an extrapolation of migrant populations for selected species.
Censuses were begun between 06:30 and 08:30. Censuses ran an average of 170.5 minutes (range 115 to 270 minutes). Census time averaged longer in spring and fall when migrants were present. Censuses were run weekly except when the area was flooded. When possible, days of high wind or rain were avoided. The census method consisted of walking slowly along the route and stopping at regular spots. All birds both seen and heard were recorded. Birds flying overhead were recorded but a notation to that effect was made.
A total of 238 censuses were conducted between February 1974 and June 1980. Of these only the 197 censuses taken from 1975 through 1979 are used in this presentation.
Data Analysis
A system of numbered weeks, starting 1–7 January was used to organize the data. All censuses taken during each week were pooled to gain the overall trend. Week 2 (8–14 Jan.) was not represented by any censuses so the figures for that week are the average of weeks 1 and 3. A three-number running average was used as a curve-smoothing device on the composite graph.
The total number of individuals (TI) and the frequency of occurrence (F) was calculated for all species seen from 1976 through 1979. The species were then ranked by a composite figure computed by multiplying TI by F (proportion of 1.0).
Species diversity (H') was calculated using the Shannon formula (Shannon and Weaver 1948). The maximum species diversity (H'max) was also calculated for each sample as was evenness (J'= H'/H'max) (Pielou 1966). Foliage height diversity (FHD) was calculated using the same formula based on presence or absence of foliage at three levels (0 to 2 m., 2 to 5 m., 5+ m.) at 700 points.
The nesting pair densities were based on spot-mapping of singing males and located nests.
Density was figured for a hypothetical km2 of similar habitat. A similar method was used for winter populations but the numbers were based on an average of all censuses from December through mid-February. The migration numbers for selected species were obtained from individual-year census data. The total number of migrants was estimated based on a three-to-four day layover for building up fat stores during migration.4
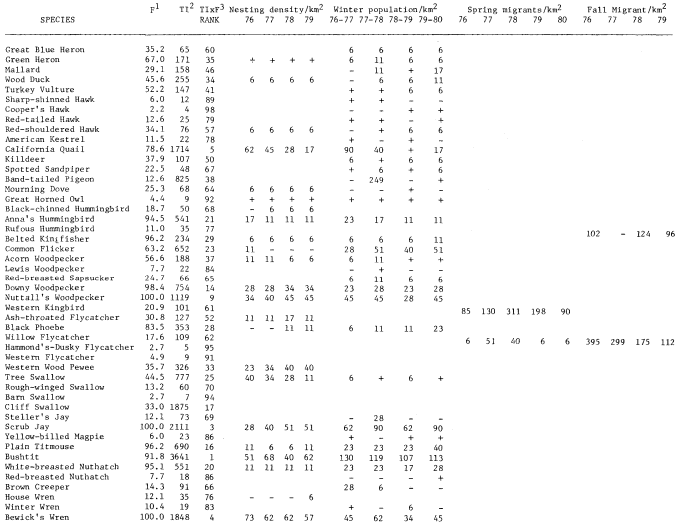
Results
A total of 147 species of birds were recorded during the censuses. The 98 species listed in table 1 occurred regularly. This table shows TI, F, Rank, and nesting, winter, and estimated migrant populations. The two forms of Yellow-rumped Warbler and Dark-eyed Junco are treated as separate taxa. Scientific names of the birds are not included to save space.
Species Diversity
Species diversity (H') is a concept which indicates both the richness and evenness of the members of a plant or animal community. It is mainly used to compare the diversity of one system or area to another.
Robert MacArthur (1972) used this formula to compare bird species diversity (BSD) with foliage height diversity (FHD) (MacArthur and MacArthur 1961). He found that FHD explained BSD better than plant species diversity or any other habitat parameter that he measured. This theory has been widely tested and has held in many cases (Balda 1975).
|
|
Species diversity of the nesting pairs on the study area was figured for each year, 1976 through 1979 (table 2). The average for the four years was 3.00 and the FHD was 1.08. All nesting BSD figures were higher than predicted using MacArthur's graph. This is not at all surprising since the study area was 17.7 ha., as compared to 2-ha. areas used by MacArthur (1972). A larger area will generally have more species nesting, and therefore will have a larger BSD. A sample of three 2-ha. plots was chosen from the 1979 nesting census data. BSD figures of 2.05, 2.40, and 2.70 were calculated for those plots. The latter two figures fit into MacArthur's linear regression, but the first figure falls well below any of his results. It is possible that the FHD was not the same for these small plots as for the whole study area.
Comparison of BSD at different seasons is also possible. When comparing the nesting and wintering population BSD, the wintering BSD was considerably higher, but shows less evenness. This is misleading due to the averaging of 6–12 census dates to obtain the wintering population figures. In order to make valid comparisons between winter and nesting populations, I chose two winter and two summer census weeks and again obtained a higher BSD on the winter samples. This was due mainly to a higher number of species, since the evenness was very similar (table 2).
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
BSD's for spring and fall census weeks were also calculated. A great deal of difference from week to week for both BSD and evenness was obtained. Overall, summer had the lowest BSD and spring, the highest. This is not surprising since the spring census weeks had the highest number of species and the summer census weeks had the lowest (table 2). When comparing weeks 16 (16–22 April) and 19 (7–13 May), the BSD was much lower during week 19 even though the species total was the same due to the large influx of Cedar Waxwings. Increased numbers of one species will always decrease the BSD.
Seasonal Changes
Figure 2 shows the changes in numbers of birds and in the total and average numbers of species from week to week, averaged over the combined 5-year period of study. The total number of species is a summation of all species found during each week for the five years of the study. The averge number of species is the average number recorded on all censuses during each week over the 5-year period. The peaks and dips in the three lines do not occur in the same places.
Total number of species (species) and average number of individuals (individuals) held relativey constant from the beginning of the year through week 5 (29 Jan.–4 Feb.). In week 6 (5–11 Feb.) and 7 (12–18 Feb.) individuals dropped while species held steady. The drop in individuals was due to a reduction in Band-tailed Pigeons in 1978. During week 8 (19–25 Feb.) individuals began to climb, reaching a peak on week 10 (5–11 Mar.). This peak was due to large increases in swallows, Audubon's Warblers, and American Goldfinches.
Species began to swing upward on week 9 (26 Feb.–4 Mar.) and rose to a peak on weeks 15 to 17 (9–29 Apr.) as spring migration was in full swing. Species dropped from weeks 17 (23–29 Apr.) through 24 (11–17 June) as the wintering species left and migrants passed through, leaving only the summer residents.
On the other hand, individuals dropped from week 10 (5–11 Mar.) through week 14 (2–8 Apr.) as the numerous Band-tailed Pigeons, Audubon's Warblers, and American Goldfinches left the area. Individuals then rose again through week 18 (30 Apr.–6 May) with migrant Cedar Waxwings, Wilson's Warblers, and Black-headed Grosbeaks making large increases. Individuals declined steadily through week 24 (11–17 June) as the migrants and winter residents left.
On week 26 (25 June–1 July) both individuals and species began to rise due to the fledging of young and influx of the first fall migrants. On week 28 (9–15 July), with species still rising, individuals began to fall as the swallows left. Individual continued to fall through week 33 (13–19 Aug.) as the common summer residents (Western Wood Pewee, Northern Oriole, and Black-headed Grosbeak) left. At the same time, species continued to rise as early migrants moved through.
Species peaked from weeks 35 to 37 (27 Aug.–16 Sept.) and then again on week 43 (22–28 Oct.) as the early and late waves of migrants passed
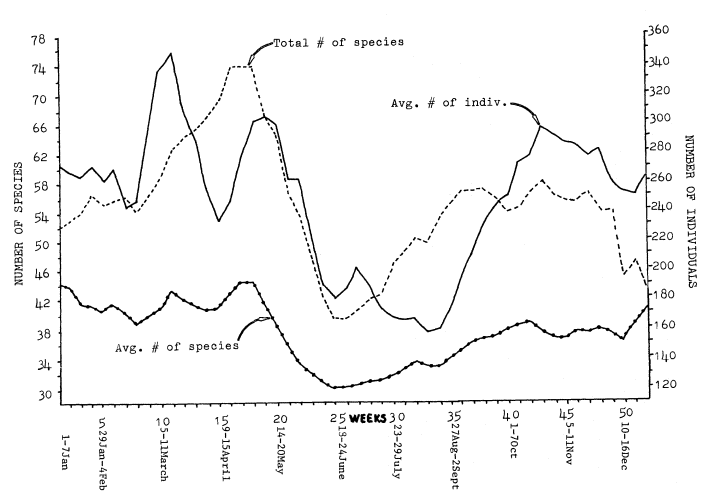
Figure 2.
Variations in numbers of species and individuals (three-week running average) for 1975 through 1979.
through. Individuals began to rise on week 34 (20–26 Aug.) and continued to go up until a peak on week 43 (22–28 Oct.) was reached. This was caused first by large numbers of Orange-crowned Warbler and Yellow Warbler, and later by an in-flux of Ruby-crowned Kinglet, Cedar Waxwing, and Audubon's Warbler. Individuals and species then dropped gradually to week 50 (10–16 Dec.).
The difference between the average number of species and total number of species was greatest during migration, and smallest during winter and summer. This was caused by a greater chance of rarities during migration, and differences in migration timing from year to year. An average of not more that 45 species was found on any week, but the chances of new species occurring on censuses during that week in succeeding years were greatest during migration.
Nesting Season
The lowland riparian nesting avifauna in central California is quite rich. A total of 83 species have been documented as nesting in the riparian areas of the Sacramento Valley in the past 100 years. Five species (Common Merganser, Spotted Sandpiper, Western Wood Pewee, Starling, and Brown-headed Cowbird) have colonized the area; and 11 species (Double-crested Cormorant, Cooper's Hawk, Bald Eagle, Long-eared Owl, Willow Flycatcher, Western Flycatcher, Purple Martin, Swainson's Thrush, Blue-gray Gnatcatcher, Bell's Vireo) have become either very rare or extirpated during that time (Gaines 1974).
Of these 83 species, 35 were found breeding on Dog Island during the study interval. An average of 29.5 species nested each year (range 27 to 31). Twenty-five species nested in all four years, two nested in three years, four nested in two years, and four nested once. Of the 10 species not found every year, five (Black-
chinned Hummingbird, Black Phoebe, House Wren, Yellow Warbler, and House Finch) colonized the area during the study. Two species (Common Flicker and American Goldfinch) became extinct as nesting species, and three species (Red-winged Blackbird, Lazuli Bunting, and Lesser Goldfinch) both colonized and became extinct as nesting species during the study.
The most abundant nesting species were the Black-headed Grosbeak, Bewick's Wren, and Bushtit. The total number of nesting birds averaged 139 pairs in 1977. In most cases, the numbers of nesting pairs of each species varied little from year to year. Notable exceptions were California Quail and Tree Swallow which declined; and Western Wood Pewee, Scrub Jay, American Robin, and Northern Oriole, which increased.
The origin of the nesting pairs was 50% residents, 13% short-distance migrants, and 37% long-distance migrants. The origin of the 35 nesting species was 44% residents, 28% short-distance migrants, and 28% long-distance migrants. Of the unstable species (i.e., those colonizing or becoming extinct), four were long-distance migrants. Only the Black Phoebe was a resident. This indicates a very unstable situation for migrants, with 45% colonizing or becoming extinct in four years.
Nesting guilds were devised on the basis of the type of nesting situation used. The avian community nesting guild composition is presented in table 3. The proportion nesting on the ground was quite low, considering the dense groundcover. This was not surprising however, since ground nesting is dangerous (Skutch 1976). The proportion of tree-hole nesters was high. This was undoubtedly a result of the large number of snags and decadent trees on the plot. These provided sites for woodpeckers to excavate their homes which in turn provided homes for the secondary cavity nesters. Many of the live trees also had natural cavities from broken-off limbs.
| |||||||||||||||||||||
Spring and Fall Migration
Each year gave a different overall impression of migration. Much of this impression came from the presence or absence of irruptive species, but much also came from the actual timing and numbers of migrants.
To gain some insight into the spring migration differences over the years, I used two criteria: 1) migration timing; and 2) actual numbers. The species accounts (Laymon 1981) and estimated spring migrant numbers (table 1) were examined for six categories: 1) record high totals; 2) record low totals; 3) record early arrival date; 4) record late arrival date; 5) record early departure date; and 6) record late departure date. The number of species that set records in any category in any year was recorded (table 4). From a birdwatcher's standpoint, 1977 was clearly the "best", since more species arrived earlier, left later, and set records for numbers. Just as clearly, 1980 was the "worst" spring, with more species arriving later, leaving earlier, and setting record low numbers. The three years 1976 through 1979 all had many early and three late departure dates, indicating a mixed situation.
Fall migration was analysed in the same manner (table 4). Late and early arrival and departure dates seemed to have little relation to the general timing of migration in fall. This was because fall migration is a much more relaxed affair. Migration timing is much less precise than in the spring. As a result, very little pattern was revealed in this portion of the analysis except that 1979 had the most early arrivals and the fewest late arrivals, and 1976 was just the reverse. The record high and low numbers revealed that 1977 had the most highs and no lows, 1976 and 1979 had the least highs and the most lows.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Winter Populations
Winter populations varied from one year to the next, much more so than breeding populations. The difference between the highest and lowest breeding years was only 14%, while the difference between the highest and lowest wintering populations was 38%. The higher totals in 1977–78 were caused almost exclusively by irruptive species such as Band-tailed Pigeon, Steller's Jay, Varied Thrush, Cedar Waxwing, Evening Grosbeak, and Pine Siskin.
Examination of a sample of 25 regularly occurring, non-irruptive species revealed that the winter of 1979–80 actually had the highest wintering population, with an average of 229 individuals per survey. The winter of 1976–77 had 160 individuals, 1977–78 had 198 and 1978–79 had 178 individuals per survey. This was still a 30% difference between the highest and lowest years. Variations in food supply, temperature, rainfall, and floodwaters must have been responsible for some of the variation in bird numbers. Much may also have been caused by nesting success and food supply on the breeding grounds.
Discussion
Yearly Changes in Numbers and Migration Timing
Numbers of migrants, both individuals and species, varied greatly from year to year. The cause of these changes was not readily evident. In some cases, the immediate weather patterns, such as rain or wind, probably grounded many migrants or kept them from leaving an area. In other cases, long-range weather patterns such as a drought, late snows, or a long, severe cold spell probably changed migration patterns. In 1977, many more than average numbers of migrants were found at Dog Island. This was a drought year in which little winter rain and no spring rain fell. It was likely that the spring migrants avoided the foothills which were parched by mid-March, and depended heavily on the lowland riparian forest for a refueling stop. It was also likely that the heavy fall migration that year was caused by the mountain springs and meadows being dry by late summer. The abundant food in the lowland riparian zone seems to be especially important to migrants during drought years.
Another factor leading to changes in numbers of migrants from year to year is nesting and wintering success. The number of birds passing north in the spring is the number which survived the winter. Winter, not breeding season, has been suggested as the season which limits bird population numbers in the temperate zone (Fretwell 1972). It is possible that destruction of riparian woodland to the south and the removal of vast areas of tropical forest may limit nesting and migrant populations here in the future. Removal of riparian woodland here may now be limiting nesting populations to the north. The birds migrating south in the fall are the sum of the surviving adults which went north to breed plus their offspring. In years of good nesting success, the number of migrants could be much higher that in poor years.
Effects of Brown-headed Cowbirds
Brown-headed Cowbirds are a recent immigrant to the Sacramento Valley of California. This bird is a parasitic species which lays its eggs in other birds' nests. The host species then incubates the eggs and raises the young, leaving the cowbird free to look for additional nests and food. The arrival of the cowbird has had a drastic effect on the native avifauna throughout California, particularly in lowland riparian systems adjacent to agricultural areas (Gaines 1974).
Had this study been done prior to the arrival of the Brown-headed Cowbird, a remarkably different avian community would probably have been present. Certain species such as the Willow Flycatcher, Western Flycatcher, Swainson's Thrush, Blue-gray Gnatcatcher, Bell's Vireo, and Warbling Vireo nested in the lowland riparian areas. None of these are now known to nest anywhere in lowland interior California. Other species such as the Yellow-billed Cuckoo, Yellow Warbler, Common Yellowthroat, and Song Sparrow still nest in the Sacramento Valley, but are now found in reduced numbers. With the exception of the Yellow-billed Cuckoo, Western Flycatcher, and Swainson's Thrush, all of the species in both groups are considered to have high to very high susceptibility to cowbird parasitism (Gaines 1974). Evidence implicating the Brown-headed Cowbird in the decline of these species is only circumstantial, since no studies were conducted during the 1940's and 1950's as the decline was occurring. However, no other theory presently explains the decline.
Of all the species which have declined or were extirpated, only the Yellow Warbler nested in the study area during the study. I believe that numbers of nesting individuals, numbers of nesting species, and species diversity were much lower in the study area due to cowbird parasitism. The species which have maintained high populations are either hole-nesters, species which reject cowbird eggs (Rothstein 1971), or species such as the Black-headed Grosbeak, which are large enough to raise a cowbird young along with their own. The small open-cup nesting species which accept cowbird eggs have all declined or disappeared completely.
Effects of the Habitat Island Nature of the Study Area
The study area is an island of riparian vegetation in a sea of farmland, oak woodland, and a river. The theories of island biogeography were developed by studying oceanic islands. These systems have higher extinction rates, lower species diversity, and fewer species than a comparable area of mainland (Diamond 1969). Distinction has been made between oceanic and habitat islands (MacArthur 1972). Habitat islands do not have as marked an effect on avian community structure, but the effect is still evident (Vuilleumier 1970).
Extinctions and colonizations of 10 nesting bird species were recorded in the four summers of the study. This represented almost a third of the nesting species, and seems like a high rate of turnover. BSD figures were at or below the predicted values using MacArthur's FHD linear
regression. Both of these findings may be related to the habitat island situation.
As more and more habitat islands are created by conversion of riparian forest to other uses, this study area will become more representative of the valley forests. If the islands are large enough, the effect will not be disastrous since the migrant riparian species in the West have been accustomed to travelling over hundreds of kilometers of desert to nest along the streams. Resident species may not fare as well, and none will fare well if the islands are too small.
An additional topic to consider regarding habitat islands is the concentration of migrants on these islands. This concentration has been documented in desert oases and on coastal islands such as the Farallons on the central California coast. It is very likely that the study area drew a much greater number of migrants because it was surrounded by miles of inhospitable habitat for forest birds than it would if a similar 18-ha. plot was found within a much more extensive stand of forest.
Values of Riparian Systems
The value of a vegetation-type can be measured in a number of ways. Some of the most common methods are bird densities, species richness, and the number of species unique to the system. These figures can be compared from system to system and a relative importance can be derived. Using American Birds breeding bird census and winter bird population studies for 1979 and 1980 I compiled table 5 which compares Dog Island results with other riparian areas and other vegetation-types in the West. No figures for migrant densities are available in the literature, so no comparisons were made.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Breeding bird densities were 24% lower at Dog Island than at 14 other riparian sites, while the number of species was slightly higher. When compared to other vegetation-types, however, the Dog Island figures were much higher, with bird densities ranging from 47% higher than chaparral to 89% higher that desert scrub. All other vegetation-types also had fewer species, ranging from 33% less in chaparral to 80% less in desert scrub. The Dog Island winter density figures are comparable to other riparian sites (5% less) and range from 57% higher than oak woodland to 89% higher than desert scrub. Winter bird species numbers were 58% higher than other riparian sites, 67% highr than oak woodland, and 85% higher that desert scrub. None of these other vegetation-types even begin to approach the riparian sites in either density or species richness.
The third category, number of species unique to the vegetation-type, is a harder one to compare. Little information has been gathered on densities of the same species in different vegetation-types, so habitat preferences are usually a matter of subjective judgment. Twenty of the regularly occurring species: Wood Duck, Common Merganser, Red-shouldered Hawk, Spotted Sandpiper, Downy Woodpecker, Black Phoebe, Willow Flycatcher, Tree Swallow, House Wren, Swainson's Thrush, Ruby-crowned Kinglet, Warbling Vireo, Yellow Warbler, Yellow-breasted Chat, Wilson's Warbler, Lazuli Bunting, Black-headed Grosbeak, American Goldfinch, Lincoln's Sparrow, and Song Sparrow are believed to have their highest densities in riparian systems. Of this group, Wood Duck, Red-shouldered Hawk, Willow Flycatcher, Swainson's Thrush, Yellow Warbler, Yellow-breasted Chat, Wilson's Warbler, and American Goldfinch are rarely found away from this vegetation-type. Is is doubtful that other vegetation-types in the West could match these figures.
In this study we have seen that a great many individuals of a great many species of birds use the Dog Island riparian system. This 17.7-ha. plot receives a great deal of avian use throughout the year. It is vital to the nesting, migrating, and wintering success of numerous individuals of many species. When viewed on a larger scale, the riparian systems of the West are vital to the maintenance of many populations, subspecies (Blue Grosbeak, Red-shouldered Hawk, and Yellow-billed Cuckoo), and, in a few cases, species of birds (Willow Flycatcher and Bell's Vireo). These systems have a high species diversity throughout the year. They are an essential link for long-distance migrants from the north and are an important wintering ground for many species.
Literature Cited
Anderson, B.W., and R.D. Ohmart. 1977. Vegetation structure and bird use in the lower Colorado Valley. p. 23–24. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: a symposium. [Tuscon, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43. 217 p. Rocky Mountain Forest and Range Experiment Station, Fort Collins Colo.
Balda, R.P. 1975. Vegetation structure and breeding bird diversity. p. 59–80. In : D.R. Smith (tech. coord.). Management of forest and range habitats for nongame birds: Proceedings of the symposium. [Tuscon, Ariz., May 6–9, 1975]. USDA Forest Service GTR-WO-1. 343 p. USDA Forest Service, Washington, D.C.
Brumley, T.D. 1976. Upper Butte Basin study 1974–75. Wildlife Management Branch administrative report 76–1. The Resources Agency, Department of Fish and Game, Sacramento, Calif.
Chase, T. Jr, and P. DeBenedictis. 1964. Middle Pacific Coast region report. Audubon Field Notes 19:71.
Dembosz, D., K. Ficket, and T. Manolis. 1972. Disturbed floodplain woodland. Amer. Birds 26:978–979.
Diamond, J.M. 1969. Avifaunal equilibria and species turnover rates on the Channel Islands of California. Nat. Acad. Sci., Proc. 64:57–63.
Fretwell, S.D. 1972. Populations in a seasonal environment. Princeton University Press, Princeton, N.J.
Gaines, D. 1973. Floodplain riparian woodland. Amer. Birds 27:995.
Gaines, D.A. 1974. A new look at the nesting riparian avifauna of the Sacramento Valley, California. Western Birds 5:61–80.
Gaines, D.A. 1977. The valley riparian forests of California: their importance to bird populations. p. 57–85. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Grinnel, J., J. Dixon, and J.M. Linsdale. 1930. Vertebrate natural history of a section of Northern California through the Lassen Peak region. University of California Publ. in Zool. 35(1).
Hehnke, M., and C.P. Stone. 1978. Values of riparian vegetation to avian populations along the Sacramento River system. p. 228–235. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Ga., December 11–13, 1978]. USDI Forest Service GTR-WO-12. 410 p. Washington, D.C.
Hurst, E., M. Hehnke, and C.C. Goude. 1980. The destruction of riparian vegetation and its impact on the avian wildlife in the Sacramento River Valley, California. Amer. Birds 34:8–12.
Ingles, L. 1950. Nesting birds of the willow-cottonwood community in California. Auk 67:325–332.
Kendeigh, S.C. 1944. Measurement of bird populations. Ecol. Monogr. 14:67–106.
Laymon, S.A. 1981. Avifauna of an island of lowland riparian woodland: Dog Island City Park, Red Bluff, California. Unpublished masters thesis, California State University, Chico.
Laymon, Stephen A. 1983. Photodocumentation of vegetation and landform change on a riparian site, 1880–1980: Dog Island, Red Bluff, California. In : R.E. Warner and K.M. Hendrix (ed.). California Riparian Systems. [University of California, Davis, September 17–19, 1981]. University of California Press, Berkeley.
MacArthur, R.H. 1972. Geographical ecology. Harper and Row, Inc. New York, New York.
MacArthur, R.H.; and J.W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.
Manolis, T. 1973. Disturbed floodplain woodland. Amer. Birds 27:994–995.
Michny, F.J., D. Boos, and F. Wernette. 1975. Riparian habitats and avian densities along the Sacramento River. The Resources Agency, Department of Fish and Game, Wildlife Management Branch administrative report 75–1.
Pielou, E.C. 1966. Species diversity and pattern diversity in the study of ecological succession. J. Theoret. Biol. 10:370–383.
Root, R.B. 1967. The niche exploitation pattern of the Blue-gray Gnatcatcher. Ecol. Monogr. 37:317–350.
Rothstein, S.I. 1971. Observation and experiment in the analysis of interactions between brood parasites and their hosts. Amer. Natur. 105:71–74.
Shannon, C.E., and W. Weaver. 1948. The mathematical theory of communication. University of Illinois Press. Urbana, Ill.
Shuford, D. 1973. Disturbed riparian stream border. Amer. Birds 27:1005.
Skutch, A.F. 1976. Parent birds and their young. University of Texas Press. Austin, Texas.
Small, A. 1974. The birds of California. Winchester Press. New York, New York.
Tangren, G. 1971. Riparian oak woodland. Amer. Birds 25:967–969.
Tangren, G. 1972. Riparian oak woodland. Amer. Birds 26:977–978.
Vuilleumier, F. 1970. Insular biogeography in continental regions. The northern Andes of South America. Amer. Nat. 104:373–388.
Winkler, D. 1973a. Riparian oak woodland. Amer. Birds 27:689.
Winkler, D. 1973b. Riparian oak woodland. Amer. Birds 27:996.
Avian Use of Marshes on the Lower Colorado River[1]
Bertin W. Anderson, Robert D. Ohmart, Julie K. Meents, and William C. Hunter[2]
Abstract.—Vegetation composition and structure was used in designating eight types of marshes along the lower Colorado River. Marsh-types that had high densities of cattail (Typhalatifolia ) and bulrush (Scirpusacutus ), supported more wading birds (including Yuma Clapper Rail [Ralluslongirostris yumanensis ]) and insectivores. Waterbirds and shorebirds favored more open marsh situations. The interface between marsh and terrestrial riparian habitat was more important to terrestrial birds than to marsh birds.
Introduction
Marshes are an important component of wildlife habitat in riparian systems. In the lower Colorado River valley, marshes frequently occur adjacent to terrestrial and aquatic habitats and may support avian species characteristic of both habitats, such as the endangered Yuma Clapper Rail (Ralluslongirostrisyumanensis ), which occurs primarily in marsh vegetation.
In the lower Colorado River valley, marshes differ in vegetation composition and structure; these differences are reflected in the avian species which inhabit them. In this paper, we describe the different types of marshes and their dominant vegetation characteristics. Marsh use by granivores (GR), shorebirds (SB), insectivores (I), waterbirds (WB), and wading birds (WA) is evaluated by marsh-type and by season.
Methods
A total of 30 transects was established in representative marsh vegetation along the Colorado River between Davis Dam, Arizona/Nevada border and the United States/Mexico International Boundary (fig. 1). Each transect was censused three times per month between May 1976 and July 1978, using a modified variable transect method (Emlen 1971; Anderson and Ohmart 1977). Results of all censuses on a transect in a particular season were averaged to obtain seasonal densities. Seasons, designated to coincide with changes in avian population, included winter (December–February), spring (March–April), summer (May–July), late summer (August–September), and fall (October–November). Bird species found in marshes were combined into groups or guilds on the basis of similarity in feeding behavior and general habitat use (table 1).
Foliage height diversity (FHD) and foliage density were measured by the method of MacArthur and MacArthur (1961). Percent dominance of each plant species was evaluated by determining the number of times each species was included in foliage density measurements. Principal components analysis (PCA) (Nie etal .) was used to determine which vegetation variables were highly intercorrelated; the PCA also provided a series of new, noncorrelated variables based on the original data. Each transect was given a score representing its relative position on the continuum represented by each of these derived variables. These scores were used in grouping transects into general marsh-types.
Results and Discussion
Vegetation
The PCA indicated that several of the vegetation variables measured were highly correlated. Cat-tail (Typha latifolia ) and bulrush (Scirpusacutus ) tended to occur together, but neither coexisted with phragmites (Phragmitesaustralis ). These three variables were combined into a derived variable; the relative amounts of these plant species are represented by the score of each transect (Fig. 2). On the
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Bertin W. Anderson is Faculty Research Associate; Robert D. Ohmart is Associate Director; Julie K. Meents is Research Biologist; William C. Hunter is Field Biologist; all are at the Center for Environmental Studies, Arizona State University, Tempe, Arizona.
Figure l.
Map of marsh transects along the lower Colorado
River.

first principal component or derived variable, positive scores indicate increasing amounts of cat-tail or bulrush; negative scores indicate increasing amounts of phragmites. Structural differences, derived from FHD measurements, indicated that cat-tail/bulrush tends to be short (below 1.5-m. height) and dense, whereas phragmites is taller (above 1.5-m. height) and more spatially heterogeneous.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The PCA also showed that trees (mostly salt cedar [Tamarixchinensis ]) and grasses tend to occur together. A transect's score on this derived variable reflects its measured density of trees and grasses (fig. 3). A negative score indicates few or no trees and grasses and a positive score indicates above-average amounts of trees and grasses.
Marsh-Type
The relative scores of the transects on these two derived variable suggested that there were basically six types of marshes on the lower Colorado River. Two additional marsh-types, an open (water) marsh and a geographically isolated marsh, were compared with the original six types.
The types were:
Type I—nearly 100% cat-tail/bulrush, small amounts of phragmites and open water;
Type II—nearly 75% cat-tail/bulrush, many trees and grasses interspersed;
Type III—about 25–50% cat-tail/bulrush, some phragmites, open water, some trees and grass;
Type IV—about 35–50% cat-tail/bulrush, many trees and grasses interspersed;
Type V—about 50–75% cat-tail/bulrush, few trees and grasses interspersed;
Type VI—nearly 100% phragmites, little open water;
Type VII—open marsh (75% water), adjacent to sparse marsh vegetation; includes sandbars and mudflats when Colorado River is low; and
Type VIII—Topock Marsh near Needles, California; vegetatively similar to Type I, but with even denser stands of bulrushes. This marsh-type was kept separate for purposes of analysis as it was censused for only five months and is geographically isolated from the rest of the study area.
Avian Occurrence in Marshes
Granivores
Highest densities of granivores occurred in most marsh-types during fall, winter, and spring (table 2). Occurrence of granivores in marshes was related to the occurrence of trees and grasses. Marsh-type II, having high occurrences of both cat-tail and bulrush and trees and grasses, would be expected to supply the largest food source for granivores. This type supported the highest density of granivores. Type IV, with average densities of cat-tail/bulrush and above-average densities of trees and grasses, and Type III, with average cat-tail/bulrush and average trees and grasses, had the second and third highest granivore densities, respectively. The lowest densities of granivores occurred in Type VI (phragmites), which had few cat-tails, bulrushes, trees, or grasses, and thus provided limited food sources. However, in Type VI, highest densities of granivores occurred during summer. Upon reviewing the species composition for this season and type, the Brown-headed Cowbird (Molothru s ater ) was the sole species involved and
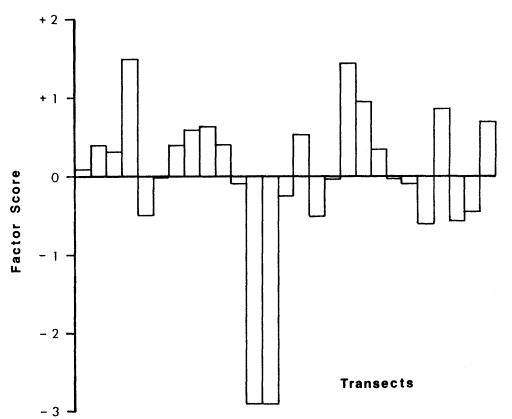
Figure 2.
Factor score of each transect on a derived variable representing the amount of cat-tail,
bulrush, and phragmites. Positive scores indicate increasing amounts of cat-tail and
bulrush; negative scores indicate increasing amounts of phragmites.
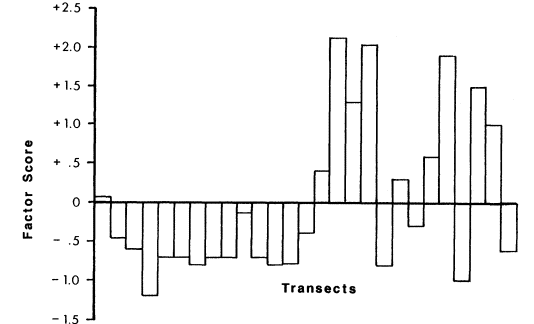
Figure 3.
Factor score of each transect on a derived variable representing density of trees and
amount of grass. Positive scores indicate above-average amounts of trees and
grasses; negative scores indicate few or no trees and grasses.
probably acted as a nest parasite rather than actually foraging on seeds.
Shorebirds
Shorebirds were most numerous during fall, winter, and spring (table 2). The highest densities occurred in open marsh (Type VII) where exposed sandbars and mudflats facilitated shorebird use. All other marsh-types had low densities; practically no shorebirds occurred in the most vegetationally dense types (I, II, V, and VI).
Insectivores
Insectivore analysis was complex due to the large number of species and the high niche separation of these species. However, the following trends were evident (table 2). Seasonally, the majority of marsh-types supported highest densities of insectivores in fall and winter. Dense and intermediately dense cat-tail/bulrush (Types I and V) did not follow this trend; highest densities of insectivores in these types occurred during summer.
Cat-tail/bulrush marshes of Types II and III had the highest insectivore densities at all seasons (especially fall and winter for Type II; see table 2). Type III had remarkably even numbers of insectivores at all seasons. These consisted mostly of Red-winged Blackbird, Song Sparrow, Common Yellowthroat, and Long-billed Marsh Wren during all seasons. Cat-tail/bulrush marshes of Type I and marshes with trees and grasses interspersed (Type V) had lower insectivore densities and followed Type III in being even at all seasons.
Though only censused during spring and summer, the very dense Type VIII marsh had very high total densities, consisting primarily of Longbilled Marsh Wren, Common Yellowthroat, and Song Sparrow. Overall for breeding insectivores, as cat-tail/bulrush became very dense, densities of Long-billed Marsh Wren, Common Yellowthroat, and Song Sparrow increased (noted in Types VIII and I). As cat-tail/bulrush approached average density, numbers of Red-winged Blackbird increased (note especially Type III; table 2). Overall, large numbers of Yellow-rumped Warbler contributed to the high fall, winter, and spring insectivore densities in most marsh-types. In open marsh (Type VI), however, high densities were accounted for by large numbers of the Water Pipit, which requires more open areas for foraging.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Waterbirds
Seasonally, waterbirds were most numerous during fall, winter, and spring in all marshtypes. Waterbirds reached their highest densities in open marsh (Type VII; table 2). Marsh-types with trees and grasses interspersed (V) and those consisting of phragmites (VI) had very low waterbird densities. The most obvious trend was for waterbirds to avoid densely vegetated areas.
However, Type VIII, a dense cat-tail/bulrush marsh, had unexpectedly high waterbird densities during spring. The American Coot was the overwhelmingly most numerous species. Though the apparent trend for denser vegetation to have
lower waterbird densities is probably realistic, it must be tempered with the realization that detectability of species in this guild is difficult due to visibility problems and the lack of highly vocal species (with the exception of the American Coot). More open vegetation would facilitate easier feeding for most species, while denser vegetation would provide cover when species in this guild were not feeding.
Wading Birds
Since the collective total for wading birds was much less than that for any other guild, alterations of marshlands could cause a severe decline in their numbers (table 2). Thus, wading birds may provide an accurate gauge for ascertaining the health of marshlands along the lower Colorado River. One species from this group, the Yuma Clapper Rail, will be analyzed in detail in the following section.
Seasonally, wading birds were most numerous in spring, summer, and late summer (table 2). Most wading birds moved out of marshland after breeding, as indicated by differences between breeding (spring and summer) and wintering seasons in most marsh-types. During summer, cattail/bulrush marshes of Types V, VIII, and I, and phragmites marshes of Type VI, in that order, were the most important types of vegetation for breeding. For wading birds collectively, the species comprising the vegetation do not appear to be as important as its density. Open marsh (Type VII) had the lowest density of wading birds. Although not taken into account in this paper, the presence or absence of tall, dead trees should be considered for any studies on habitat use for breeding herons and cormorants, as they use the dead trees for nesting platforms.
Yuma Clapper Rail
The Yuma Clapper Rail is endemic to the marshes of the lower Colorado River from Needles, California south to the Colorado Delta of Mexico, including immediate drainage areas (Salton Sea, Gila River, and Salt River). An analysis of marsh use of the Yuma Clapper Rail was necessary because of its close relationship with marshes and because of the threatened status of this subspecies. The analysis covered only spring, summer, and late summer, as the majority of the Colorado River population is migratory (Smith 1975).
Clapper Rail densities were compared for each of the defined marsh-types by month (table 3). The importance of the different marsh-types was determined by ranking. During each month, the marsh-type with the highest rail density received the highest score; the marsh-type with the lowest density received the lowest score. Scores for each marsh-type were then added across months and compared directly.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Type V was by far the marsh-type most highly used by the rails. Type I ranked one-half as high as Type V, but was nearly double the rank of Types II, III, VI, and VIII. Marsh-types with trees and grasses interspersed (Type IV) and very open marshes (Type VII) consistently ranked low. Of interest, Type VI (mostly phragmites) had relatively high ranking during spring when rails arrive to breed, and Type VII (above-average density of cat-tail/bulrush and above-average density of trees and grasses) had very high ranking during late summer when rails dispersed from the breeding areas. But during summer breeding, these marsh-types were not used extensively by rails. Even though rails appear to select certain marsh-types for actual breeding, nonbreeding marginal areas may still be important.
Conclusions
Among the six avian categories or guilds considered, five occurred in significantly greater densities in at least one marsh-type, relative to the density in the marsh-type with the lowest density. Thus, the first conclusion is that the species within these guilds have definite habitat preferences. Among the six guilds, peak populations were reached in four different marsh-types. Thus, the second conclusion is that to maximize densities and diversities of birds using marsh as habitat, it is necessasry to have a variety of marsh-types available.
A third conclusion is that among the marshtypes studied, dense stands of phragmites supported the lowest avian densities and diversities. But even this marsh-type was potentially valuable to the Yuma Clapper Rail in the prebreeding season and to granivorous species, especially in winter, spring, and late summer.
Unrestricted vehicular traffic has been found to reduce the use of marsh and open-water areas by waterfowl (Ohmart and Anderson 1978; Anderson and Ohmart 1981). One simple means of habitat improvement would be to close roads adjacent to at least some marshes during periods of avian peak use, i.e., winter.
While man has created a large portion of the existing marshes along the lower Colorado River (Ohmart etal . 1975), we hasten to point out that channelization and riprapping activities, in general, reduce the quality of marshes (ibid .). Riprapping and channelization activities should be undertaken only if the creation of marsh habitat is part of a mitigation proposal.
Immediate action must be taken to support the mesquite (Prosopisglandulosa ) skeletons in Topock Marsh to ensure a nesting area for the heron and cormorant rookery, which may well be the largest remaining rookery along the lower Colorado River.
Channelization projects usually result in destruction of sandbars and mudflats. Our final conclusion is that when these operations occur, an assertive stance must be taken to ensure that mitigation includes plans for creation of new sandbars and mudflats in the general areas.
Acknowledgments
We would like to thank Helen Wood for presenting this paper. Susan M. Cook and Jane R. Durham provided editorial assistance and Marcelett Ector typed the various drafts of the manuscript. Cindy D. Zisner typed the final draft. Judy Huff, Janet Jackson, Ann Kasprzyk, and Helen Wood constructed the tables and graphs. Rodney H. Ohmart and Elaine Hassinger prepared the final figures. We wish to give special acknowledgment to Alton Higgins who was instrumental in conducting the field work. This study was supported by USDI Bureau of Reclamation Contract Number 7-07-30-V0009.
Literature Cited
Anderson, B.W., and R.D. Ohmart. 1977. Climatological and physical characteristics affecting avian population estimates in Southwestern riparian communities. p. 193–200. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitats: a symposium. [Tucson, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Anderson, B.W., and R.D. Ohmart. 1981. Vegetation management. Final report to USDI Bureau of Reclamation, Boulder City, Nevada. In preparation.
Emlen, J.T. 1971. Population densities of birds derived from transect counts. Auk 88:323–342.
MacArthur, R.H., and J.W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.
Nie, N.H., J.G. Jenkins, K. Steinbrenner, D.H. Bent. 1975. Statistical package for the social sciences. McGraw-Hill, Inc., New York, N.Y. 675 p.
Ohmart, R.D., and B.W. Anderson. 1979. Wildlife use values of wetlands in the arid southwestern United States. p. 278–295. In : P.E. Greeson, J.R. Clark, and J.E. Clark (ed.). Wetland functions and values: the state of our understanding. Proceedings of the national symposium on wetlands. [Minneapolis, Minn., November 1978]. American Water Resources Association, Minneapolis, Minn. 674 p.
Ohmart, R.D., W.O. Deason, and S.J. Freeland. 1975. Dynamics of marshland formation and succession along the lower Colorado River and their importance and management problems as related to wildlife in the arid Southwest. p. 240–251. In : Transactions of the 40th North American Wildlife and Natural Resources Conference. Wildlife Management Institute, Washington, D.C.
Smith, P.M. 1975. Habitat requirements and observations on the Clapper Rail (Ralluslongirostrisyumanensis ). Unpublished M.S. Thesis, Arizona State University, Tempe, Ariz. 35 p.
Riparian Forest as Habitat for the Least Bell's Vireo[1]
M. Violet Gray and James M. Greaves[2]
Abstract.—Data are presented on vegetation structure in Least Bell's Vireo (Vireobellii pusillus ) breeding habitats. A discussion of vegetation and several other factors relevant to breeding success of this endangered species is presented. Recommendations are made for actions to ensure the future of the subspecies as a breeding inhabitant of riparian forests.
Introduction
The purposes of this paper are to present data and a discussion of the use of riparian forest by the Least Bell's Vireo (Vireobelliipusillus ), its breeding success over two seasons, and an assessment of the features in the habitat which are important to breeding success. The data are the results of a study initiated in 1979 and continued through the 1980 and 1981 breeding seasons in Gibraltar Reservoir watershed in Santa Barbara County, California.
The Least Bell's Vireo (referred to here as vireo) has undergone a rapid decline in California in both numbers and breeding range over the last several decades. Once present as far north as Chico, California (Cogswell 1958), and numerous enough to be considered a common summer resident (Grinnell 1928), the species' present known northern breeding range limit is Santa Barbara County. Only about 200 pairs are thought to still breed in the state (Wertz 1981). The species is considered to be heavily parasitized by the Brown-headed Cowbird (Molothrusater ) (McCaskie 1975), with a rate as high as 58% (Goldwasser 1978). The cowbird has been blamed for the vireo's decline in California (Tate 1981). However, we believe that the cowbird problem is merely a symptom of a more crucial problem—the loss or degradation of habitat suitable to sustain healthy breeding riparian bird communities.
Over the past 100 years, the riparian zones of many rivers have been heavily grazed; major cities have diverted water supplies from water-courses supplying riparian areas; flood control districts have channelized, denuded of vegetation, and often concrete-lined, streambeds; and overuse of water for irrigation has depleted the water tables in many of the state's richest river valleys. As the many prime riparian forests have disappeared, the vireos have disappeared as well, and now survive mostly in remote areas where riparian systems are relatively unaltered; or they attempt to breed (often with no success) in areas where the vegetation has been highly degraded. In these latter areas, cowbirds are often abundant and thus have significant negative effects on the reproductive capacities of most breeding song birds, including the vireo. As the most obvious agent of breeding failure, cowbirds receive an undue portion of the blame.
The riparian vegetation of the Gibraltar Reservoir study area was relatively undisturbed and supported the largest known population of the California subspecies of the vireo—about 60 pairs. By providing data on the use of this unaltered vegetation by the vireo, we hope to aid in establishing guidelines for habitat rehabilitation throughout the bird's breeding range in the state.
Description of Study Area
The study area comprises 243 ha. (600 ac.) and consists of four sections, each approximately 1.6 km. (1 mi.) in length, containing similar degrees of forestation. The Santa Ynez River, for 1.6 km. (1 mi.) to the southeast and 1.6 km. (1 mi.) west of its confluence with Mono Creek, makes up half the study area. Southeast of the Mono Creek confluence, the river extends for about 11.3 km. (7 mi.) upstream to Juncal Dam (Jameson Lake). Only the lower 1.6 km. of this reach of river is used regularly by the vireo, although a few have been found nesting in thickets above the study area. West of the confluence is the silted-in eastern third of Gibraltar Reservoir (on the Santa Ynez River) which, over the
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] M. Violet Gray is a field biologist presently contracted to the USDA Forest Service, Santa Barbara District, Calif. James M. Greaves is a biologist presently contracted to the USDA Forest Service, Santa Barbara District, Calif., and Curatorial Assistant, Museum of Natural History, Santa Barbara, Calif.
past 40 years, has developed into a broad silt floodplain with a narrow creek channel bordered by lush riparian forest. This area contains the habitat for approximately one-third to one-half of the vireo population (hereafter Gibraltar Reservoir population).
Mono Creek joins the Santa Ynez River at the east end of the reservoir and has approximately the same flow as the river. The creek is obstructed about 1.6 km. (1 mi.) above the confluence by a debris dam built in the 1930s. Above this dam is the broad bowl of Mono Debris Basin, fed by Mono Creek, Indian Creek, and Little Caliente Creek. One-quarter of the vireo population breeds in the 1.3-sq. km. (0.5-sq. mi.) basin.
The study area is surrounded by rugged mountains with slopes often exceeding 40°. The slopes are very dry; many have open rocky patches that support little or no vegetation. Chaparral communities dominate the majority of the slopes, except where intense fires have recently burned, promoting the growth of grasslands.
There are several plant communities that can be classified as riparian vegetation which are not necessarily dominated by willow/cottonwood forest, and which are associated with vireo breeding. The Gibraltar/Mono study area contains at least four of these communities: oak woodland; willow/cottonwood forest; shrubby thicket; and dry wash. Each of these communities is dependent upon the regular flow of water through canyons, and its stability is determined by the river's flow and flood levels.
Oak Woodland
This is the most permanent of the four communities. It occupies the drier sites in the study area and is transitional between the mesic willow/cottonwood forest and the xeric chaparral-scrub communities. Coast live oak (Quercusagrifolia ), squaw bush (Rhustrilobata ), poison oak (R . diversiloba ), elderberry (Sambucusmexicana ), and snowberry (Symphoricarposmollis ) are common elements of this community. There is a great deal of intrusion by species more typical of chaparral communities, such as Ceanothus spp., chaparral honeysuckle (Lonicera johnstonii ), and basin sagebrush (Artemisia tridentata ).
This community is the only one of the four in which grasses predominate as the primary herbaceous groundcover. Introduced species of brome grass (Bromus spp.) and wild oats (Avena fatua), as well as native mugwort (Artemisiadouglasiana ) form the major groundcover species. In openings between the oaks or in less shaded areas beneath oaks, thickets of shrubby species have developed. These thickets seldom reach 1.8 m. (6 ft.) in height and are usually limited to about 1.2 m. (4 ft). Wild blackberry (Rubusursinus ) and California wild rose (Rosacalifornica ) are occasional components of these areas and seem to occupy the ecotone between oak woodland and willow/cottonwood forest as well as being regular associates of the latter community. Tall shrubs and young trees do not generally form a continuous layer here. Young oaks, buck brush (Ceanothuscuneatus ), red heart (C . spinosus ), elderberrry, and coyote bush (Baccharispilularis ) appear as salients from the more continuous lower layer of shrubs. Thus, there is an open aspect to this community which is accentuated by the sparsely foliated habit of many of the dominant taxa. Mature oaks in this area are commonly 15.2 m. (50 ft.) high, with particularly large individuals exceeding 21.3 m. (70 ft.).
Willow/Cottonwood Forest
This community occurs in two basic forms. The first is that of a dense-canopied forest with little or no vegetation below the canopy level. The second form has the same canopy structure, but is also rich in shrubby and/or herbaceous undergrowth. The canopy generally consists of a mixture of Fremont cottonwood (Populus fremontii ) and red willow (Salixlaevigata ), often growing to heights of 9.1–18.3 m. (30–60 ft.). The majority of the willows are not much greater than 229 mm. (9 in.) diameter-at-breast-height (DBH), many having little girth despite their great heights. In a few areas, most notably in the Mono Debris Basin, both willow and cottonwood are found with trunk diameters approaching 1.2 m. (4 ft.). The cottonwood grows in smaller numbers, but to slightly greater size, scattered among the willow. Where cottonwood exist to the exclusion of willow, it takes the form of either evenly distributed gallery forest or large trees growing in grove-like clusters of six to 12 trees, in an open woodland arrangement.
Another common species, arroyo willow (S . lasiolepis ), does not generally grow to sufficient size to become a part of the forest canopy layer, but some rather slender trees mixed in with red willow have been found with heights up to 13.7 m. (45 ft.). Other taxa found in this lower canopy, but to a lesser degree, are: western sycamore (Platanus racemosa ), black cottonwood (Populustrichocarpa ), black willow (S . gooddingii ), yellow willow (S . lasiandra ), and white alder (Alnus rhombifolia ).
The low shrub layer consists partially of taxa which are herbaceous, such as mugwort, but which have the habit of growing in shrub-like clumps and which function (with respect to several nesting bird species) as low shrubs. This layer also contains shrubs which do not attain heights greater than 1.2 m. (4 ft.) or which are at a stage of intermediate growth. Nearly all the plants from the other layers, except those forbs and grasses which usually do not attain heights of 0.6 m. (2 ft.), can be found in the low shrub layer.
The structure of the high shrub layer differs from that of the low shrub layer in that the lower 0.6–1.2 m. (2–4 ft.) of the plants in the
high shrub layer are generally devoid of dense foliage. Unless it occurs in conjunction with the low shrub layer or groundcover, the absence of foliage at the lower level in the high shrub layer makes high shrubs of little use to nesting vireos in the study population. All of the willow species contribute to the high shrub layer, either as young or adult trees or in the form of shoots from downed larger trees. Narrowleaf willow (Salixexigua ) is a major element of this layer, along with wild blackberry, California wild rose, snowberry, and seep-willow (Baccharisglutinosa ). Immature trees of all the above-mentioned species are occasional components of the high shrub layer.
The groundcover in these forested areas is generally composed of only one or two species, with white sweet-clover (Melilotusalbus ) and mugwort predominating. Many moisture-requiring forbs, such as peony (Paeoniacalifornica ), speedwell (Veronicaanagallis-aquatica ), and hedge-nettle (Stachysalbens ), were present at several survey sites.
Shrubby Thicket
The thicket is the simplest vegetative community in the study area. It generally possesses neither a canopy nor much groundcover and often is composed solely of one plant species, the most common of which are narrowleaf willow and black willow. The thicket presents an outer aspect which appears to be a solid wall from the ground to the top of the stand, often reaching 6.1 m. (20 ft.) when mature. The wall of vegetation is formed by a combination of low and high shrub layers growing at the edge of the thicket. The interior, which is usually the most mature section, is seldom as dense as the exterior. This community usually occupies loose sandy or rocky ground which is not stable enough to support the larger forest types.
Dry Wash
These areas are generally useless to vireos for breeding, except when thickets establish themselves and are sufficiently large to provide foraging habitat or nest sites. On occasion a tree or small thicket has grown in such a way in a dry wash to withstand the heavy flooding of winter runoff, and vireo nests have been found in such sites. These areas produce cover very slowly because they are heavily washed and scoured every year. There is some encroachment at the edges by thickets of narrowleaf or arroyo willow, which seem to combine rapid maturing rates with strong tolerance for unstable sandy or gravelly soils.
There is some groundcover in the dry washes each year, depending on how heavily scoured they have been and how high the previous winter runoff was. Wild mustard (Brassicacampestris ), white sweet-clover, and cocklebur (Xanthium strumarium ) usually dominate, with some chaparral and oak woodland plants interspersed throughout.
Large piles of debris have created other major open spaces in the study area. These are found scattered within riparian woodlands and consist of matter ranging from large cottonwood and willow trunks to mats of dry foliage mixed with silt. They seldom support much vegetation other than introduced weeds such as curly dock (Rumexcrispus ) or heliotrope (Heliotropiumcurassavicum ).
Methods
Before 1979, no in-depth breeding data existed for the Least Bell's Vireo, and there was little information about its habitat requirements. In 1979, the authors began a study of the Gibraltar Reservoir population, banding and color-banding young birds as groundwork for a long-term population study. In 1980 and 1981, we continued these studies and began gathering data on vegetation at individual nest sites and along transect lines established in 243 ha. (600 ac.) of forest comprising the study area.
Because the vireo was the focus of this study, intense field work was required to locate nest sites, determine the number of pairs, and conduct the banding studies. We developed methods in the field for the search-and-find part of the project, walking throughout the habitat, following natural courses as much as possible, listening and looking for vireos. Whenever a male or a pair was encountered, we searched its territory until a nest was located or it was determined that it was not nesting. Adults were mistnetted when practicable, preferably prior to incubation of eggs, and young were banded between the ages of 6 and 10 days (10 to 12 days nestling life). We monitored each nest for success or failure and cause of failure; measured parameters for possible reasons for success; and recorded numbers of eggs, hatchings, and fledgings.
The vegetation study consisted of two parts: a habitat description taken from observation in the field (see DESCRIPTION OF STUDY AREA), and a quantitative analysis of transect data and nestsite surveys. To determine in detail what features of the vegetation are important to vireos in the study area, we surveyed sample plots at 0.04-ha. (0.1-ac.) circles around nest sites (James 1971, modified), and at 161 sites along 11 transect lines arranged across streamcourses. Transect plots were spaced 10 m. apart, each plot alternating to the right or left of the line and having an area of 3 m. square. All data were gathered over each square meter within each transect plot. These transects covered a total length of 2,093 m. Data collected at all sites included: frequency or absence of cover by class; frequency of plant species; and density of shrubs. Four cover-classes were used: groundcover (less than 60 cm. height), low shrub (600–120 cm.), high shrub (greater than 120 cm.), and canopy (greater than 6 m. and stems greater than 7.6 cm. [3 in.] DBH).
Results and Discussion
Vireo Breeding
Arrival and Territory
Most males and some females arrived in the study area in mid-April. Males initiated nest building immediately upon their arrival, with the females apparently deciding whether to use the nest as is, complete it, or build elsewhere. The remainder of the population drifted in over the following two to four weeks. Territories ranged in size from 0.2 ha. to 1.6 ha. (0.5 ac. to 4.0 ac.), the size usually dependent on availability of food and water, and sufficient vegetation for at least several nest sites.
Nest Location
Nest heights averaged 64 cm. (range 33–133 cm.) aboveground, with successful nests about 8 cm. lower than failed nests. Most nests were placed near or at the edges of thickets, or at the edges of woods and open fields or washes, in low shrub or upper groundcover levels. A few nests were placed in full sun adjacent to, but not protected by, abundant cover. Some nests were built in galleried forests and were usually placed in tangles of groundcover or low shrubs at the bases of large trees. Most nests were hidden well behind a screen of leaves, limbs, or dead material, or combinations of live and dead material. However, high contrast (light and shadow moving across the nest during the day) seems to have contributed more to nesting success than either frequency of occurrence or density of vegetation around the nests. With only a few exceptions, nests were suspended: a) from forks or parallel stems or twigs of live woody material under 5 mm. in diameter; b) from dead twigs of willow or cottonwood; or c) from dead stems of mugwort or white sweet-clover under a cover of live material, usually mugwort.
Nesting and Production
Every known vireo pair nested at least once, usually exploring several sites before settling on a suitable one. Most pairs built a least two nests; as many as six nests were located in one territory. Several pairs built only two nests, successfully rearing young in both, and one pair built a third nest after its second fledging (table 1).
Several successful pairs were able to raise only one nestling in 1981 from each of two successful nests in their territories, in contrast to the normal three or four young raised per nest by most of the population. In 1979 and 1980, the only one-chick nests were the result of cowbird activity, or were begun late in the season when food and water stress would be expected to be higher. Thus, the lower fledgling number per pair, in spite of a higher nesting success rate, may have been the result of a higher-than-normal amount of nonpredation nestling mortality.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Feeding Patterns
As the 1981 season progressed, vireos were seen foraging farther into the chaparral, often traveling several hundred yards to obtain food for their nestlings. This pattern of activity was apparent to a greater extent in 1979 and 1981, indicating the possibility of cyclic food shortages. We and others observed that the vireo does not leave its brood exposed during the 14- to 16-day incubation period (Hensley 1950). During the 1981 season, however, many nests were left unattended for long periods of time with neither adult present nearby. We suspect the reason for this was a combination of extremely high temperatures and low food availability which necessitated the absence from the nest for long periods by both adults because neither was able to obtain enough food for its own sustenance during its turn away from the nest. High daytime temperatures aided in incubation, but humidity
was insufficiently maintained around the eggs, and, as a result, many freshly hatched young were weakened and unable to survive the rigors of life outside the egg.
Departure from the Study Area
Adult birds departed the study area as early as mid-July, and all were gone by the beginning of September. Most young birds left or were not seen in their natal territories after the fourth week from fledging. However, in 1981 many first-clutch young were present during second breeding attempts and disappeared from their territories at the time the second brood was ready to fledge, often found over 1.6 km. (1 mi.) away in another vireo's territory. Generally, females and young left the study area before males.
Use of Habitat
The vireo behaves both as a specialist and a generalist within separate and particular elements of the habitat. One major constraint is that the species nests solely in California's southern riparian zones. Since these vireos often raise two clutches of four young each and food is therefore a primary consideration during the breeding season, they would be expected to breed in areas of high humidity and insect productivity such as the riparian zones.
The Vireo as Specialist
As mentioned, nests are placed in a narrow range of heights, rarely higher than 1.8 m. (6 ft.). Since such higher placements are generally reactions to radical environmental disturbances (e.g. sudden removal of lower vegetation layers),4 they do not appear to reflect a trend toward use of diverse nesting levels.
Nests are usually near the edge of a thicket, or, if in a small shrub or thicket, they may be in the middle. Usually all nests are within 1 m. (3 ft.) of an opening. This arrangement allows at least one unobstructed flight path to the nest, easy visibility of predators, and good foliage cover for nest safety. For small birds incapable of much actual defense at the nest (Best and Stauffer 1980), all of these factors contribute to successful nesting attempts. The branching arrangement of the immediately supporting nest substrate is usually a horizontal or down-sloping fork, or horizontally parallel stems.
The Vireo as Generalist
Within the constraints of the above-stated requisite elements, the vireo is a generalist. Nest substrate, in terms of plant species, seems to correlate fairly closely to those plant taxa available and abundant, with a few notable exceptions. California wild rose and coast live oak were both used more often than they were encountered in the study area transect lines. This may be because they associate closely with other species (willows and mugwort, respectively) which are abundant and widespread (table 2). The vireo is also a generalist in its selection of plant life forms, using forbs, shrubs, and trees for both nest support and cover. It generalizes more broadly in nest cover, often preferring large-leaved plants, or mixtures of leaf size-classes.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Shrub density at nest sites seems to correspond to density throughout the forest, indicating that the vireos do not seek out any unusually dense stand of vegetation for nesting. There is an extremely broad range of densities both at nest sites and at transect plots, so density is apparently not of importance to the bird in nest-site selection (table 3).
| ||||||||||||||||||
Cover-type frequencies were generally similar in both samplings (nest site and transect plots) (table 4), so presence of a particular cover-type such as canopy does not appear to be a consideration in nest site selection.
| ||||||||||||||||||||||
Although no data were recorded indicating time allotments in any one level or community, the vireos were observed foraging in all levels and communities in and adjacent to the study area during all parts of the breeding season. However, they seemed to spend a major portion of their time gleaning insects from leaves in the willow and cottonwood forests at and below the middle levels of the canopies. Early to midseason feeding activity was limited generally to areas near nest sites, while later season feeding encompassed all communities, including a good portion of time in adjacent chaparral.
Recommendations
These recommendations are made for general management of Least Bell's Vireo breeding habitat and for rehabilitation of historical or degraded sites.
1. Prohibit removal of groundcover and low shrubs.
2. If non-native plant species are removed, they should be replaced immediately (within one season) with a suitable native of comparable form.
3. Limit or eliminate recreation and grazing or other agricultural uses in sensitive riparian systems or critical habitat.
4. Replanting of denuded areas should include a diversity of species and should include species which provide all cover-type layers.
Acknowledgments
This research was sponsored by USDA Forest Service, Santa Barbara District, Calif.; California Department of Fish and Game, Non-game Management Branch; Santa Barbara Audubon Society; USDI Fish and Wildlife Service, Office of Endangered Species. We thank Alan Craig, Maeton Freel, and Sanford Wilbur for their personal aid in obtaining funding. We also thank contributing Audubon Society chapters: Altical, Los Angeles, Morro Coast, and Santa Barbara.
Literature Cited
Best, Louis B., and Dean F. Stauffer. 1980. Factors affecting nesting success in riparian bird communities. Condor 82:149–158.
Cogswell, H.L. 1958. Middle Pacific Region. Audubon Field Notes 12:379–384.
Goldwasser, Sharon. 1978. Distribution, reproductive success and impact of nest parasitism by Brown-headed Cowbirds on Least Bell's Vireos. 27 p. California Department of Fish and Game, Sacramento.
Goldwasser, Sharon, David Gaines and Sanford R. Wilbur. 1980. The Least Bell's Vireo in California: a de facto endangered species. Am. Birds 34:742–745.
Grinnell, J. 1928. A distributional summation of the ornithology of Lower California. Univ. California Publ. Zool. 32(1):1–300.
Hensley, M.M. 1950. Notes on the breeding behavior of the Bell's Vireo. Auk 67:243–244.
James, F.C. 1971. Ordinations of habitat relationships among breeding birds. Wilson Bull. 83:215–236.
McCaskie, G. 1975. Southern Pacific Coast Region. Am. Birds 29:1029–1036.
Tate, James, Jr. 1981. The blue list for 1981. Am. Birds 35:3–10.
Wertz, Paul. 1981. Update on endangered species. Outdoor California 42:1–6.
Importance of Riparian Systems to Nesting Swainson's Hawks in the Central Valley of California[1]
Ronald W. Schlorff and Peter H. Bloom[2]
Abstract.—Once a common breeding bird of the Central Valley and elsewhere in California, the Swainson's Hawk (Buteoswainsoni ) has experienced a severe population decline due to, among other things, loss of the riparian systems that provided trees for nest sites. The pattern of land use prevalent over the past 130 years has reduced riparian systems of the Central Valley to a tiny fraction of their former extent. It will be necessary to maintain and restore stands of vegetation in riparian systems if the Swainson's Hawk is to continue as a breeding species in the Central Valley.
Introduction
The Swainson's Hawk (Buteoswainsoni ) (fig. 1) was once assumed to be so common in California that most historical reporters felt it did not warrant special mention (Bloom 1980). It was variously described as the most abundant hawk in the southern Transverse Ranges (Sharp 1902) and common in spring, summer, and fall from the foothills to the ocean (Willett 1912). Willet (ibid .) also reported a population of Swainson's Hawks breeding on Santa Catalina Island. Historic accounts provide some qualitative data on the former abundance of the species, but no quantitative studies of Swainson's Hawk population size or breeding distribution in California were published from the late nineteenth and early twentieth centuries. Thus, today, it is difficult to assess the status when quantitative data on the historic population are lacking. Fortunately, however, early records of the egg-collectors do provide useful information on historic distribution of the species.

Figure 1.
Adult male dark-phase Swainson's Hawk marked with colored
leg band and USDI Fish and Wildlife Service band.
Bloom (1980) used a combination of factors to arrive at a historic statewide nesting population estimate of between 4,300 and 17,100 pairs of Swainson's Hawks. The method involved taking observed nesting densities in relatively undisturbed habitat at the present and extrapolating backward to include all suitable nesting and foraging habitat assumed to be available to the species historically. By comparing estimated historic population levels with that observed today, it is evident that the Swainson's Hawk has suffered a population decline of enormous proportions, perhaps greater than a 90% statewide decline since 1900 (Bloom ibid .).
Comparison of historic and present range reveals the species is now absent from many regions of California where it once was a common breeding bird (figs. 2 and 3). Historic population estimates for the Central Valley ranged from 1,656 to 6,624 pairs. During 1979, a breeding population of 280 pairs was estimated inhabiting the area from 40 km. (25 mi.) south of Fresno in the San Joaquin Valley north to the vicinity of Chico in the Sacramento Valley.
Valley populations of Swainson's Hawks frequently nested in valley oaks (Quercuslob -
[1] Paper presented at the California Riparian Systems Conference. [University of Califogrnia, Davis, September 17–19, 1981].
[2] Ronald W. Schlorff is Wildlife Biologist, Nongame Wildlife Program, California Department of Fish and Game, Sacramento, Calif. Peter H. Bloom is Raptor Biologist, Santa Cruz Predatory Bird Research Group, University of California, Santa Cruz.
Figure 2.
Historic range of the Swainson's Hawk
in California. This is the approximate
range during the late nineteenth and
early twentieth centuries (shaded area).
Figure 3.
Current range of the Swainson's Hawk in California.
This range was delineated from results of surveys
conducted during 1979–81 (shaded area).
ata ) and Fremont cottonwoods (Populus fremontii ) historically and at the present time. Often, trees chosen as nest sites are within 90 m. (100 yd.) of a major Valley stream, creek, or slough in the riparian zone. Today, remnants of riparian vegetation exist along portions of the Sacramento, American, and Feather Rivers and their tributaries in the Sacramento Valley, and along the San Joaquin River and tributaries in the San Joaquin Valley (fig. 4) (Thompson 1977; Conard etal . 1977). Additional riparian vegetation suitable for nesting Swainson's Hawks may be found along the many miles of creeks, sloughs, and canals that crisscross the agricultural and grazing fields of the Central Valley. These habitats often support the only trees in a local agricutural area, and therefore are vital to Swainson's Hawks for nest sites.
The 1979 Swainson's Hawk study (Bloom 1980) provided the basis for subsequent investigations into the species' dependence on riparian systems for nesting in the Central Valley. Monitoring of nest sites in 1980 and 1981 confirmed the initial indication of the importance of riparian systems to Swainson's Hawks. Unfortunately, a pattern of destruction and degradation has characterized much of man's relationship with this vegetation-type in the Central Valley and elsewhere (Thompson 1977; Smith 1977). Smith (ibid .) estimated that about 314,000 ha. (775,000 ac.) of riparian vegetation existed in the Central Valley in 1850. By 1977, only 4,900 ha. (12,000 ac.)—1.5%—remained. This loss is continuing and there are plans to remove more vegetation along the Sacramento River and other riparian ecosystems in order to effect "bank protection" and for "flood control" (Jannssen 1976). If species such as Swainson's Hawks are to survive, the trend of destruction must be reversed.
Methods
Study Area
During 1979, the entire state was surveyed for nesting Swainson's Hawks. For purposes of this paper, the habitat relationships of the species in the Central Valley were investigated. An area corresponding to the Valley floor and extending from 40 km. (25 mi.) south of Fresno to the vicinity of Chico, approximately 47,700 km2 (18,400 mi2 ) constituted the study area (fig. 3). There are still significant riparian systems available to nesting Swainson's Hawks within this area. A 219 km. (135 mi.) section of the Sacramento River, from Colusa south to the Sacramento Delta, served as a riparian study area and was surveyed to determine nesting density and habitat quality. Additional riparian areas consisting of large and small streams, sloughs, and canals also provided study areas.
Habitat Description
Historic and present Central Valley riparian systems have been described in detail (Conrad etal . 1977; Michny etal . 1975; Roberts etal . 1977; Thompson 1977). Wildlife studies document habitat relationships of the

Figure 4.
Typical Swainson's Hawk nesting habitat in a Central
Valley riparian system. Similar stands of vegetation
exist along portions of the Sacramento River.
various species dependent on riparian systems (Brumley 1976; Gaines 1977).
The riparian zone of major Valley streams has been described as a continuum of plant communities following the topographic line from the stream channel through the low and high terrace deposits of the floodplain (Conard etal . 1977). Cottonwoods, oaks, sycamores (Platanusracemosa ), and large willow trees (Salix sp.) form the dominant overstory vegetation in the zones most important to Swainson's Hawks. Cottonwoods typically dominate the lower terrace deposits closer to the stream bank while sycamores and valley oaks are found on higher terrace deposits and cut banks along the outside of meanders (Conard etal . 1977). Several species of woody and herbaceous plants comprise the understory in this riparian forest. Historically, and to a lesser extent today, a native grassland community, including oat (Avena sp.), brome-grass (Bromus sp.), ryegrass (Elymus sp.), barley (Hordeum sp.), and ryegrass (Lolium sp.) provided foraging habitat for Swainson's Hawks beyond the valley oak component of the riparian system.
Survey Techniques
Techniques used in the 1979 Swainson's Hawk survey are described in Bloom (1980). Subsequent surveys in 1980 and 1981 were made employing these same methods for automobile surveys of riparian vegetation along major watercourses and smaller streams and sloughs. Systematic search of areas known or suspected to support Swainson's Hawks provided nesting data for the two years following the 1979 survey.
In 1981, a 219 km. (136 mi.) portion of the Sacramento River was surveyed by motorboat to record sightings, territories, and nest sites of Swainson's Hawks. The boat survey was conducted over three days with approximately 64 to 80 km. (40 to 50 mi.) surveyed per day. In addition to recording data on the hawks, notes and maps were made of habitat availability and quality along the survey route. These maps and notes were compared against past and recent aerial photographs made in the study area. Automobile surveys in areas near to the boat survey route were made to ensure more complete coverage, and the automobile surveys better facilitated assessments of foraging areas near the river.
An analysis of habitat characteristics was made from nesting survey reports made during 1979–81. Data were pooled for the three years and information was summarized on the proximity of territories to riparian systems, and tree species and heights chosen for nesting by Swainson's Hawks.
Results
Review of 1979 Survey
Providing details on habitat association was not a primary objective of the 1979 Swainson's Hawk survey but it was an important secondary goal. The first evidence of the importance of riparian systems to nesting Swainson's Hawks was apparent during the portion of the study involving the Central Valley. Elsewhere in the State, the Klamath Basin for instance, Swainson's Hawks do not appear to be dependent on riparian vegetation because nest trees were located away from the few streams in the area (Bloom 1980).
In the Central Valley, there are also many suitable nest trees available away from riparian systems; however, their continued existence is threatened in many cases. Where these trees grow in the midst of agriculture fields, regeneration is severely hampered due to land use activities. Sometimes livestock grazing prevents regeneration, and always there is the threat of removal if the space trees occupy and the water they use is required in order to expand agricultural activities. Therefore, it is important to preserve nesting habitat in riparian systems. As noted during the 1979 survey, severe destruction of habitat already has occurred in the riparian systems and is continuing at an alarming rate.
A majority of all nests and territories recorded for the Central Valley were close to riparian systems (figs. 5, 6, and 7). Nests were found most often in cottonwoods and oaks (table 1). Cottonwoods were considered an important nest tree because they provide an excellent nest site at the present and, due to rapid growth rate, will be an ideal species for restoration work. Oaks also are important; however, regeneration time will be much longer in areas where oaks are to be restored.
Tree height was an important parameter affecting nesting, and Swainson's Hawks appeared to choose relatively tall trees for nest sites.
Tree heights ranged from 6–30 m. (20–100 ft.) with a mean of 12 m. (41 ft.) (N = 36). Nests were built near the tops of oaks and cottonwoods that provided shade for the nest and also afforded a good view of the surrounding terrain.
Subsequent Surveys
During 1980 and 1981, results of surveys continued to show the importance of riparian systems to Swainson's Hawks (figs. 5, 6, and 7). Occasional nests were found a mile or more away from riparian zones, but a greater proportion but a greater proportion
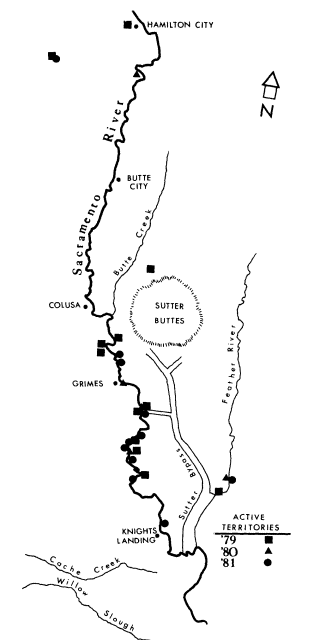
Figure 5.
Location of Swainson's Hawk nest sites in the Central
Valley of California, northern section. Data derived
from surveys conducted during 1979–81.
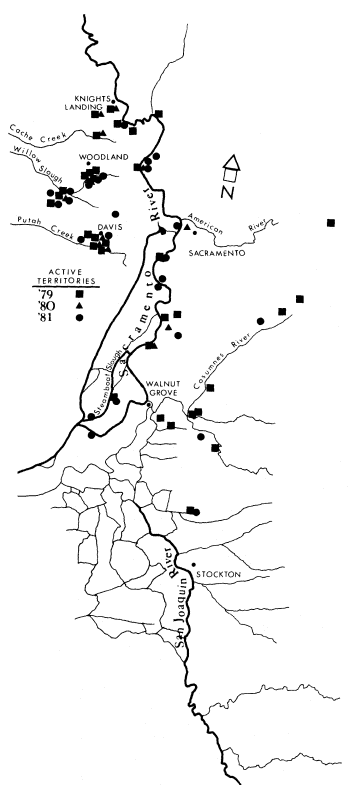
Figure 6.
Location of Swainson's Hawk nest sites in the
Central Valley of California, middle section.
Data derived from surveys during 1979–81.
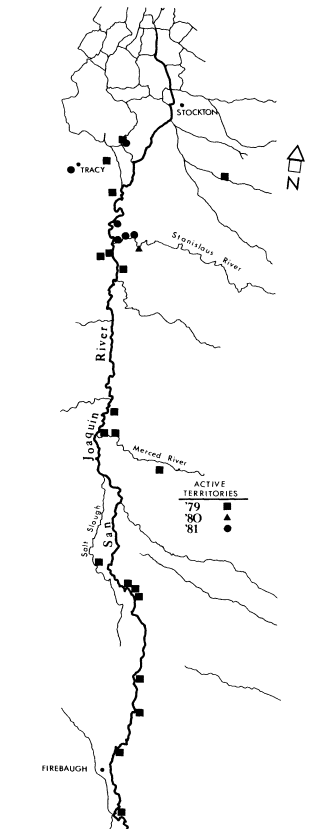
Figure 7.
Location of Swainson's Hawk nest sites in the
Central Valley of California, southern section. Data
derived from surveys conducted during 1979–81.
| |||||||||||||||||||||
were within streamside vegetation (table 2). There is a built-in bias concerning the search for nest trees. Since most suitable trees are in riparian systems in the Valley, it naturally follows that these areas supported the most nesting of Swainson's Hawks. Results of the 1981 boat survey revealed extensive habitat loss along both banks of a 219 km. (136 mi.) stretch of the Sacramento River from Colusa to the Sacramento Delta. It was estimated that 85% of the stream bank was either entirely devoid of vegetation or the vegetation that did remain was unsuitable for nesting Swainson's Hawks.
Where suitable habitat did remain and prerequisites for nesting seemed to be met, Swainson's Hawks often were found (fig. 4). Results indicate that the hawks nested in or near the riparian zone at a frequency of one pair per 13.7 river km. (8.5 river mi.); this is considerbly lower than Bloom's estimate of one territory per 6.4 km. (4.0 mi.) of suitable nesting habitat for Central Valley watercourses (Bloom 1980). The greatest nesting frequency was recorded in 1979 when three active territories were located along a 1.1 km. (0.7 mi.) stretch of Willow Slough in Yolo County. This indicates the abundance of Swainson's Hawks possible given optimum habitat conditions.
| |||||||||||||||||||||||||||
Discussion
It has been noted that the Swainson's Hawk, although not an obligate riparian species, nevertheless is closely associated with riparian systems in certain situations (Bent 1963). The Central Valley appears to be such a situation. Elsewhere in North America, the species is often found nesting in juniper (Juniperus sp.)/sagebrush (Artemesia sp.) and prairie habitats of the Great Basin (Olendorff 1975; Dunkle 1977; Fitzner 1978). The northern California population of Swainson's Hawks exists in such habitat in the Klamath Basin (Bloom 1980).
Land use patterns prevalent over the past 125 years have destroyed the once vast riparian forests that existed. Where gallery forests 6.4- to 8.1-km. (4- to 5-mi.) wide existed there are now kilometers of riprapped banks denuded of even brushy vegetation. The loss came about as a result of man's activities to obtain fuel, building material, and farmland in an earlier era and, in recent decades, to provide for agricultural expansion, "bank protection" and "flood control" (Smith 1977; Gaines 1977; Sands 1978).
Corresponding with the time of riparian vegetation loss in the Central Valley, the Swainson's Hawk population declined catastrophically. The decline has been on the order of 80–90% of the estimated historic level, a decline equal in severity to that suffered by the endangered American Peregrine Falcon (Falcoperegrinusanatum ) (Bloom 1980; Herman etal . 1970). Indeed, the Swainon's Hawk is a candidate for inclusion on the State's list of endangered species.
Besides habitat loss, other factors, such as problems relating to their migration route and wintering grounds, may have contributed to the species' decline in California. However, it seems clear that the tremendous loss of the riparian vegetation, that today appears so important to the Swainson's Hawk in the Central Valley, must have had a correspondingly tremendous impact on the population.
The solution to the problem of a declining wildlife population is never easy. There are many factors over which we may have little or no control. However, it is clear that we do have an opportunity to affect one critical factor in the Central Valley. A vigorous program to maintain and enhance remaining riparian systems may go a long way to help restore Swainson's Hawks to a more viable status. By maintaining a large and healthy Swainson's Hawk population in the Central Valley, which will serve as the core from which some of the former range may once again be repopulated, it is hoped we can mitigate for those mortality factors which we cannot control (i.e. problems on the wintering grounds in South America). Maintaining suitable nesting habitat in the Central Valley may be the key to saving the Swainson's Hawk from extirpation in California.
Literature Cited
Bent, A.C. 1963. Life histories of North American birds of prey: Part one. Dover Publications, New York, New York. p. 222–236.
Bloom, P.H. 1980. The status of the Swainson's Hawk in California, 1979. Federal Aid in Wildlife Restoration, Project W-54-R-12, Nongame Wildl. Invest. Job Final Report 11–8.0. 24 p. + appendix.
Brumley, T.D. 1976. Upper Butte Basin study 1974–1975. California Department of Fish and Game, Wildlife Management Branch, Administrative report no. 76–1. 30 p. + appendix.
Conard, S.G., R.L. MacDonald, and R.F. Holland. 1977. Riparian vegetation and flora of the Sacramento Valley. p. 47–55. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Dunkle, S.W. 1977. Swainson's Hawks on the Laramie Plains, Wyoming. Auk 94:65–71.
Fitzner, R.E. 1978. Behavioral ecology of the Swainson's Hawk (Buteoswainsoni ) in Washington. Ph.D. Thesis. Washington State University, Pullman. 194 p.
Gaines, D.A. 1977. The valley riparian forests of California: their importance to bird populations. p. 57–85. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Herman, S.G., M.N. Kirven, and R.W. Risebrough. 1970. Peregrine Falcon decline in California. Audubon Field Notes 24(4):609–613.
Jannssen, R. 1976. Army Corps of Engineers projects on the Sacramento River. In : Gaines, D.A. Abstracts from the conference on the riparian forests of the Sacramento Valley. Davis Audubon Society. 25 p.
Michny, F.J., D. Boos, and F. Wernette. 1975. Riparian habitats and avian densities along the Sacramento River. California Resources Agency, Department of Fish and Game. Administrative Report No. 75–1. March 1975. 42 p.
Olendorff, R.R. 1975. Population status of large raptors in northwestern Colorado, 1970–1972. In : J.R. Murphy, C.M. White, and B.E. Harrell (ed.). Population status of raptors. Raptor Res. Rep. No. 3:185–205.
Roberts, W., J.G. Howe, and J. Major. 1977. A survey of riparian forest flora and fauna in California. p. 3–19. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Sands, A. 1978. Public involvement in riparian habitat protection, a California case history. p. 216–227. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Georgia, Dec. 11–13, 1978]. USDA Forest Service GTR-WO-12. USDA Forest Service, Washington, D.C. 410 p.
Sharp, C.S. 1902. Nesting of Swainson's Hawk. Condor 4:116–118.
Smith, F. 1977. Short review of the status of riparian forests in California. p. 1–2. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Thompson, K. 1977. Riparian forests of the Sacramento Valley, California. p. 35–38. In : A. Sands (ed.). Riparian forests in California: Their ecology and conservation. Institute of Ecology Pub. 15. University of California, Davis. 122 p.
Willett, G. 1912. Birds of the Pacific Slope of Southern California. Pac. Coast Avifauna 7:47.
Sensitivity of Riparian Birds to Habitat Loss[1]
Julie K. Meents, Bertin W. Anderson, and Robert D. Ohmart[2]
Abstract.—The extent and composition of riparian plant communities in the lower Colorado River valley have historically been altered, primarily by man. Some of these communities are disappearing (cottonwood and mesquite), and others are expanding (salt cedar and arrowweed). We examined the avian community associated with riparian vegetation and identified avian habitat specialists. Nearly all of these specialists are concentrated in cottonwood/willow or honey mesquite communities. Salt cedar generally supported no avian species with narrow habitat breadth.
Introduction
Riparian vegetation of the Southwest desert serves as habitat islands for many species of wildlife. The lush vegetation and water along riparian corridors is generally surrounded by relatively sparse, dry uplands that are not suitable for most riparian birds. Within riparian vegetation there are various plant communities dominated by one or more plant species. Along the lower Colorado River, we have identified six major communities. The plant communities vary in their areal extent and distribution throughout the valley (Anderson and Ohmart 1976). Some, such as the cottonwood/willow and mesquite communities, have become increasingly restricted to small areas over the past 100 years because of harvesting for fuel, clearing for agriculture and urban development, wildfires, and changes in water cycles (Ohmart etal . 1977). Many of the factors that have led to the decline of native communities have promoted the establishment and spread of the exotic salt cedar; salt cedar-dominated vegetation is one of the few communities that is stable or expanding in the lower Colorado River valley.
Many avian species found in riparian vegetation are ubiquitous, but some species occur in only one or a few plant communities. Restriction to a few plant communities may limit an avian population's persistence in an area if the favored plant communities are rare or require a very long time to redevelop after disturbance. Dependence on a limited variety of plant communities may be a particular problem if an avian species always occurs in low densities. Such species typically have low tolerance for even short-term habitat loss or alteration.
In this paper we examine the avian communities of the lower Colorado River valley and identify those species that may be considered plant community specialists. The current status and future potential of the plant communities in which these specialists are found are also discussed.
Methods
Major vegetation-types were defined according to the dominant tree or shrub (table 1). Cottonwood and willow trees frequently co-occurred, so they were combined as one vegetation-type. Salt cedar occurred with other tree species throughout the study area, but was considered a separate type only where it was relatively homogeneous. Salt cedar/honey mesquite was considered a separate vegetation-type because there were extensive areas that supported an approximately equal mix of the two species. Each vegetation-type was further divided into as many as six structural types based on the vertical distribution of the foliage. Structural Type I was tallest and was multi-layered (fig. 1). Vegetation structural-types are referred to as communities throughout this paper.
The study area included the riparian floodplain of the Colorado River, from Davis Dam on the Arizona/Nevada border south to the Mexican border. Two to 11 transects, each 800–1,600 m. long, were established in stands of each community roughly in proportion to their occurrence in the valley. Birds were censused three times per
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Julie K. Meents is Field Biologist; Bertin W. Anderson is Faculty Research Associate; Robert D. Ohmart is Associate Director; all are at the Center for Environmental Studies, Arizona State University, Tempe, Arizona.
| ||||||||||||||||
month between May 1976 and July 1979 along these transects, using a modified variable transect method (Emlen 1971, 1977; Anderson etal . 1977). Seasonal averages of avian densities were calculated from all censuses taken in summer (May–July) and winter (December–February). Bird species considered in this paper include those whose occurrence and relative density were fairly predictable and stable between years. This group included most residents, as well as summer and winter visitors. Scientific names of all birds considered are listed in table 2.
Habitat breadth (HB) was calculated using the equation:
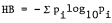
where p. equals the proportion of a species' population occurring in community i. Evenness (J) is the percent of maximum HB:
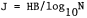
where N equals the number of communities and reflects how evenly the members of a species are distributed throughout the available habitat. We arbitrarily chose J = 0.75 as a dividing point between habitat specialists (J < 0.75) and habitat generalists (J > 0.75), based on results of censuses and our knowledge of each species.
Figure 1.
Illustrations of relative densities of trees and understory shrubs in each of
the vegetation-structural types. See Anderson et al . (1977) for details.
Results and Discussion
Summer Birds
Eighteen bird species were considered common breeding species in the lower Colorado River valley; seven of these are resident throughout the year. The majority (71%; table 3) of the summering species were found to be relatively evenly distributed throughout the plant communities and were considered habitat generalists. Because these species usually occur in all community-types, it seems likely that loss of one or a few plant communities would not constitute a serious threat to their respective populations.
Species with relatively narrow habitat breadth (habitat specialists) are discussed in greater detail below.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Gila Woodpecker
The Gila Woodpecker is fairly common in southern Arizona, where it nests primarily in saguaro cactus (Carnegiea gigantea ) (Phillips etal . 1964). In the lower Colorado River valley, the Gila Woodpecker is most frequent in cottonwood/willow and Type III honey mesquite communities (table 3). This species is a primary cavity nester and, because of its relatively large size, requires large trees for hole excavation and as a foraging substrate (Brush etal .[3] ).
Wied Crested Flycatcher
The population of Wied Crested Flycatcher breeding in the lower Colorado River valley is peripheral to the species' major distribution in Mexico. This flycatcher is primarily found in cottonwood/willow and mesquite communities (table 3). It is a secondary cavity nester and requires excavations or holes in fairly large trees.[3]
Summer Tanager
The Summer Tanager in the lower Colorado River region is also peripheral to the species' major range of distribution. It is mostly found in cottonwood/willow Types I–IV and salt cedar Type I (table 3); these are all communities with a large amount of vegetation at higher levels.
Yellow-billed Cuckoo
The Yellow-billed Cuckoo in the Southwest is limited to cottonwood, willow, or mesquite along rivers (Phillips etal . 1964). The birds in the Colorado River valley are concentrated in cottonwood/willow Types I–III, but are also found occasionally in honey mesquite Type III and screwbean mesquite Type II (table 3).
Bell's Vireo
The Bell's Vireo had a very narrow habitat breadth as well as a very low total population (table 3). Birds were generally found only in Type III or IV vegetation, but there was no association with a particular tree species. Although the Bell's Vireo probably has specific vegetation structure requirements, these do not appear to be limiting its population size, because Types III and IV vegetation are the most widespread types in the valley. Alternatively, the population appears to be most influenced by the rate of cowbird parasitism (Phillips etal . 1964; Serena 1981). The Brown-headed Cowbird (Molothrus ater ) has become increasingly common in the valley as agriculture has expanded; this has led to increased parasitism of such birds as the Bell's Vireo. In riparian systems of the Rio Grande River, Bell's Vireo occurs both in greater numbers and in a wider variety of communities (Anderson and Ohmart unpublished data); cowbird populations are also lower and agricultural land is less widespread in this area. Thus it appears that Bell's Vireo in the lower Colorado River valley is probably not primarily limited by habitat availability. The Yellow Warbler (Dendroica petechia ) has also been scarce or absent as a breeding bird in recent years in the lower Colorado River valley because of cowbird parasitism (Phillips etal . 1964).
Winter Birds
Five of 18 wintering bird species examined were habitat specialists (table 4).
[3] Brush, T., B.W. Anderson, and R.D. Ohmart. Habitat selection related to resource availability among cavity-nesting birds. 15 p. Unpublished manuscript.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Gila Woodpecker
The Gila Woodpecker was the only resident species that was a habitat specialist; this was consistent in both seasons. As in summer, it was primarily limited to cottonwood/willow and honey mesquite Type III communities (table 4), where large trees suitable for cavity excavation and foraging were available.
Phainopepla
The Phainopepla was concentrated in honey mesquite communities and had a very narrow habitat breadth (table 4). Anderson and Ohmart (1978) have shown that the association between Phainopepla and mesquite is secondary; the birds rely heavily on the berries from mistletoe (Phoradendroncalifornicum ) that infests mesquite trees. Mistletoe berries provide a major source of food for Phainopepla during winter and early in the breeding season.
Sage Sparrow
The Sage Sparrow had the narrowest habitat breadth of the wintering species considered and was confined to honey mesquite communities (table 4). This relationship also appears to be secondary, because the Sage Sparrow is usually found associated with inkweed (Suaedatorreyana ), a shrub that grows in patches in mesquite communities (Meents etal .[4] ).
Brown Creeper
The Brown Creeper occurred exclusively in cottonwood/willow communities with tall, mature trees. This species is on the verge of extinction in the valley.
Black Phoebe
The Black Phoebe seldom occurred outside of cottonwood/willow communities. Peak densities occurred in stands that were moderately open in the layer >7 m. Usually there was water in the immediate vicinity.
Conclusions
Most habitat specialists in the lower Colorado River valley are found in cottonwood/willow or honey mesquite communities. These plant communities were formerly widespread in the valley (Ohmart etal . 1977), and it is probable that the bird species that now have narrow habitat breadths were once much more abundant. Salt cedar is not favored by most bird species in the area (Cohan etal . 1978; Anderson and Ohmart 1981a). The avian species with narrow habitat breadth that we studied were rarely found in salt cedar vegetation or occurred only in the most mature stands. The first conclusion is that the continued spread of salt cedar will have a negative effect on these species. Arrowweed is another plant community that appears to be stable or spreading because it is common in disturbed areas; it is also little used by most bird species. Some bird species in the lower Colorado River valley have benefited or at least partially adapted to recent changes in proportions and composition of the local plant communities (Conine etal . 1978; Anderson and Ohmart 1981a, b). This is also demonstrated by those species with habitat breadths approaching 100% of maximum. But populations of at least six of the species discussed here are potentially threatened by the continuous decline or removal of cottonwood, willow, and mesquite vegetation. The second conclusion is that efforts should be made to revegetate areas with these plant species.
Because food resources for birds appear to be superabundant in summer (Anderson and Ohmart unpublished data), it is likely that summer habitat specialists select vegetation primarily on the basis of nesting requirements. Wintering birds are more likely to be limited by food resources, and their habitat selection probably reflects food availability. Phainopepla feeds on mistletoe which is found almost exclusively in honey mesquite communities. There is also indirect evidence that the Sage Sparrow's association with inkweed is food related (Meents etal .[4] ). The third conclusion is that habitat requirements of wintering as well as breeding populations must be studied. Secondary species of vegetation known to have high value, such as mistletoe and inkweed, should be planted in revegetation efforts.
It should be noted that avian species that require large, mature trees (Gila Woodpecker, Wied Crested Flycatcher) may be especially sensitive to habitat loss because even artificial replacement of vegetation may require many years. The Bell's Vireo and other species are hurt directly as riparian vegetation is converted to agricultural uses. Agricultural areas provide habitat for species such as the Brown-headed Cowbird. As this nest parasite increases in numbers, the number of fledged young of parasitized species, such as the Bell's Vireo, will decrease further.
Acknowledgments
Numerous field biologists helped collect the data. Susan M. Cook and Jane R. Durham provided editorial assistance, and Cindy D. Zisner typed the final copy. Brian Woodridge prepared the figure. We thank Ron Haywood for presenting the paper. We want to thank all of the above people for their tremendous dedication and help. This study was supported by USDI Bureau of Reclamation Contract No. 7-07-30-V0009.
[4] Meents, J.K., B.W. Anderson, and R.D. Ohmart. In review. Vegetation relationships and food of Sage Sparrows wintering in honey mesquite habitat.
Literature Cited
Anderson, B.W., R.W. Engel-Wilson, D.G. Wells, and R.D. Ohmart. 1977. Ecological study of southwestern riparian habitats: techniques and data applicability. p. 146–155. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: A symposium. [Tucson, Arizona, July 9, 1977.] USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Anderson, B.W., and R.D. Ohmart. 1976. Vegetation type maps of the lower Colorado River from Davis Dam to the southerly International Boundary. 4 p. plus 23 maps. USDI Bureau of Reclamation, Lower Colorado Region, Boulder City, Nevada.
Anderson, B.W., and R.D. Ohmart. 1978. Phainopepla utilization of honey mesquite forests in the Colorado River valley. Condor 80:334–338.
Anderson, B.W., and R.D. Ohmart. 1981a. Vegetation management final report. USDI Bureau of Reclamation, Boulder City, Nevada. In preparation.
Anderson, B.W., and R.D. Ohmart. 1981b. Agricultural final report. USDI Bureau of Reclamation, Boulder City, Nevada. In preparation.
Cohan, D.R., B.W. Anderson, and R.D. Ohmart. 1978. Avian population responses to salt cedar along the lower Colorado River. p. 371–381. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Georgia, December 11–13, 1978.] USDA Forest Service GTR-WO-12, Washington, D.C. 410 p.
Conine, K.H., B.W. Anderson, R.D. Ohmart, and J.F. Drake. 1978. Responses of riparian species to agricultural habitat conversions. p. 248–262. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Georgia, December 11–13, 1978.] USDA Forest Service GTR-WO-12, Washington, D.C. 410 p.
Emlen, J.T. 1971. Population densities of birds derived from transect counts. Auk 88:323–342.
Emlen, J.T. 1977. Estimating breeding bird densities from transect counts. Auk 94:455–468.
Ohmart, R.D., W.O. Deason, and C. Burke. 1977. A riparian case history: the Colorado River. p. 35–47. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitat: A symposium. [Tucson, Arizona, July 9, 1977.] USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Phillips, A., J. Marshall, and G. Monson. 1964. The birds of Arizona. 212 p. University of Arizona Press, Tucson.
Serena, M. 1981. Distribution, habitat requirements, and reproductive success of Arizona Bell's Vireo (Vireobelliiarizonae ) along the lower Colorado River. California Department of Fish and Game, Nongame Wildlife Investigations Final Report.
Avian Use of Revegetated Riparian Zones[1]
Bertin W. Anderson and Robert D. Ohmart[2]
Abstract.—We reintroduced native riparian vegetation on three plots located along the lower Colorado River. Vegetational growth and avian colonization of the areas occurred rapidly. Fremont cottonwood (Populusfremontii ), willow (Salixgooddingii ), and quail bush (Atriplexlentiformis ) were important to increased avian use. Selectively clearing salt cedar (Tamarix chinensis ) and leaving native vegetation, including dead trees, resulted in further enhancement.
Introduction
We have studied riparian vegetation and its associated wildlife along the lower Colorado River since 1972. These studies were conducted in two phases. In the first phase, we classified the riparian vegetation (Anderson and Ohmart 1976) and determined the avian densities and diversities associated with each vegetation-type. The second phase of our work included the reintroduction of native plant species and the replacement of exotic vegetation with native vegetation. In this report we address three major questions: 1) at what rates do birds begin using a revegetated area?; 2) what combinations of native vegetation have the greatest enhancement value?; and 3) how much wildlife is lost when salt cedar is cleared? We will not provide complete answers here, but we do provide significant data pertaining to these questions.
Methods
Our revegetation efforts were conducted on three sites ranging in size from 10–30 ha. Two sites were established on barren dredge-spoil. The third initially consisted of a mixed stand of salt cedar and willow. The latter site was selectively cleared, leaving the willow intact.
An avian census transect was established on each site. Transects 1,050 m. long were censused three times each month using a modified version (Anderson and Ohmart 1977) of the line transect technique developed by Emlen (1971). Foliage density estimates were obtained with the board technique (MacArthur and MacArthur 1961). The number of trees greater than 3 m. tall was ascertained. The number of shrubs was estimated by counting all shrubs within 15 m. of either side of the transect.
Foliage height diversity (FHD) was calculated using – S pi lnpi , where pi was the proportion of foliage density in one of four layers. Layers used were 0 to 0.6 m., 1.5 to 3.0 m., 4.6 to 6.1 m., and ³ 7.6 m. Each transect was divided into units 150 m. long. An index to horizontal foliage diversity (patchiness) for each of the four layers was calculated as the variance in foliage density at each level. Total patchiness was the sum of the variances (patchiness) at each of the four layers (Anderson etal . 1978). Quantified vegetation variables found on the revegetation sites were compared to the average for all of the riparian vegetation-types we recognized in the lower Colorado River Valley.
Avian use of revegetated areas was determined by comparing avian densities and diversities on revegetated areas with the average for all the riparian vegetation-types we recognized along the lower Colorado River. Vegetation development and avian use were expressed in standard units: the mean for riparian vegetation assumed a value of zero; values greater than the mean had positive values; and those less than the mean had negative values. In our study of the riparian vegetation, transects were established on all of the recognized vegetation-types approximately in proportion to their occurrence in the Valley.
The effect of clearing salt cedar was determined by comparing average densities and diversities found over a 3-year period (108 censuses) on the third plot, which was subsequently cleared. We studied the area for two years after it was cleared (72 censuses) before
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Bertin W. Anderson is Faculty Research Associate, Center for Environmental Studies, Arizona State University, Tempe, Arizona. Robert D. Ohmart is Associate Director, Center for Environmental Studies, Arizona State University, Tempe, Arizona.
it was revegetated. We recognized five seasons on the basis of climatic changes and major changes in the avifauna. According to our criteria, March and April were spring; May, June, and July—summer; August and September—late summer; October and November—fall; and December, January, and February—winter. Thus the preclearing study encompassed 15 seasons and the postclearing study conducted spanned 10 seasons.
Results
The vegetation and avian variables studied are listed in table 1. Averages for vegetation variables found for all riparian vegetation are presented in table 2, and averages for avian variables are presented in table 3.
Foliage Development of Revegetated Sites
Means of the standardized variable (habitat quality index) were not significantly different between any two of the revegetation sites or from the mean for riparian vegetation in the Colorado River Valley (table 4). This is misleading because similar means can result from fundamentally different structural configurations. For example, total foliage densities were similar on the revegetation sites, and these were similar to the mean for riparian vegetation. However, on the revegetated sites this was due to above-average foliage density at low levels, whereas in the riparian vegetation in general there was above-average density at higher levels. The FHD was above average before clearing but was markedly below average on the revegetation sites. The dredge-spoil sites had above-average numbers of cottonwood and willow trees greater than 3 m. by the end of the second year. The cleared site had above-average numbers of cottonwood and willow trees from the beginning as a result of the selective clearing. Shrubs were above average by the end of the first year on all three revegetation sites. The number of shrubs was below average on the cleared site before clearing.
| ||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The herbaceous vegetation on the revegetated plots, classified as shrubs, was composed of different species, and the total number of shrubs varied among the plots (table 5). The east dredge-spoil site had the smallest amount of herbaceous vegetation; this was almost exclusively Russian thistle (Salsolaiberica ). On the west dredge-spoil site the total density of herbaceous vegetation was about 50% greater than on the east dredge-spoil site; there was more smotherweed (Bassiahyssopifolia ) and quail bush on the west dredge-spoil site. The greatest number of shrubs occurred on the cleared site; smotherweed, inkweed (Suaedatorreyana ), and quail bush were the numerically dominant species.
| ||||||||||||||||||||||||||||||||||||||||||
Bird Use of Revegetated Sites
On the dredge-spoil sites, bird use, expressed as standard units, was above the average for riparian systems in the Colorado River Valley for almost all categories in fall, winter, and spring, but below average at other seasons (table 6). Insectivorous birds and doves were the last bird groups to colonize revegetated sites. Granivorous bird species reached above-average densities earlier than other groups and densities of insectivores tended to remain lower than average over the first two years. Bird species richness and densities were greater in winter and fall than at other seasons. Granivorous birds reached greatest densities on the west dredge-spoil and cleared sites. In general, birds used the cleared site to a greater extent a year earlier than the dredge-spoil sites. Summer densities were below the riparian average on all sites.
Impact of Clearing Salt Cedar
Clearing salt cedar resulted in removal of more than 90% of the vegetation present on the site. Cottonwood, willow, screwbean mesquite (Prosopispubescens ), and dead trees remained; their aggregate density was about 30 per ha. The site remained in this condition for two full years. The only change during that time was invasion by a few annuals, inkweed, and quail bush. By fall of the second year there was a total of 15 inkweed and quail bush on the site. Leveling, installing the irrigation system, and augering holes were activities largely responsible for keeping vegetation from developing on the site before planting.
In spite of the removal of most of the vegetation, total species richness, species richness, and densities of passerine granivores, and densities of doves were all significantly higher (P < 0.05) than before clearing (table 7). The density of insectivorous species dropped significantly for visiting insectivores but not for permanent resident birds.
Discussion
Avian colonization of sites revegetated with native vegetation was rapid, reaching average or above-average densities and diversities in less than two years. Revegetation efforts on dredge-spoil sites progressed more slowly than on the site from which salt cedar had been cleared, and the pattern of avian use reflected this. There are at least three reasons for this. First, because selective clearing was done, there were mature trees on the cleared site from the beginning. On the dredge-spoil sites there was less than one mature tree per hectare when planting began. It was not until the second year that an equal number of trees reached a height of 3 m. Second, on the cleared site we made an extensive effort to establish inkweed and quail bush. The avian data collected after clearing, but before shrubs were planted, revealed that these two shrubs are very important in attracting birds.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
We have presented additional data elsewhere in support of this claim (Anderson etal . 1978). Herbaceous vegetation, primarily Russian thistle and smotherweed, developed rapidly on the dredge-spoil sites. Granivorous, but not insectivorous, bird species were attracted to this vegetation. Insectivorous birds were attracted to the cleared site to a greater extent than to the dredge-spoil sites where quail bush was abundant. Finally, several trees with large dead snags were among the trees left on the cleared site, consequently attracting greater numbers of those bird species which use dead limbs and cavities. In areas where trees were planted. but no dead trees were present, it will be years before large dead snags develop naturally. This disadvantage might be overcome by putting up nest boxes and by girdling a few 4-year-old trees. Girdling, of course, will kill the tree unless it is done in the winter, when new growth will develop from suckers at the base of the tree.
Because avian densities and diversities increased after clearing, but before planting, it seems that merely clearing salt cedar results in some enhancement. We have presented additional data elsewhere indicating the negative value of salt cedar to most riparian birds (Anderson etal . 1977; Cohan etal . 1978). After clearing, the areas became more attractive to birds such as the Lesser Nighthawk (Chordeilesacutipennis ) and Loggerhead Shrike (Lanius ludovicianus ), which typically occur in more open areas. Granivorous bird species were probably attracted to the area after clearing because of the annuals and the few scattered shrubs which developed. Annuals and shrubs were totally absent before clearing. Doves were more abundant during the breeding season before the area was cleared, but they were less abundant at other seasons. Some species of permanent resident birds decreased significantly in numbers after clearing (e.g. Abert Towhee (Pipilo aberto)), but several species (e.g. Roadrunner (Geococcyz californianus )) came into the area (Anderson and Ohmart 1981).
If quail bush, inkweed, cottonwood, and willow are planted, frugivorous species, such as the Phainopepla (Phainopeplanitens ), will not use the area. This species, and frugivorous
species in general, exist mainly on the fruit of mistletoe (Phoradendroncalifornicum ) (Anderson and Ohmart 1978). In the Colorado River Valley mistletoe infests honey mesquite (Prosopisglandulosa ) to a far greater extent than any other vegetation. If honey mesquite is not planted in a revegetated area, little use by frugivores can be anticipated.
Colorado River water is perhaps the most sought-after water on the North American continent. Riparian vegetation uses water through evapotranspiration; water which, theoretically, could otherwise be used by man. In addition, there is concern that the vegetation in the floodplain of the lower Colorado River could obstruct the passage of floodflows. The numeriically dominant species of vegetation along the lower Colorado River is the exotic salt cedar (Anderson and Ohmart 1976). In this report we have shown that, at least under certain circumstances, a great deal of salt cedar can be cleared without having a negative impact on birds in general. It follows that if salt cedar is replaced with native riparian shrubs and trees, avian populations as well as the native riparian vegetation can be enhanced while total foliage density decreases. Theoretically, the clearing would therefore result in water salvage through reduced evapotranspiration and an increased capacity for the floodplain to carry floodwater.
Conclusions
We conclude that avian populations can be rapidly enhanced by revegetating riparian zones with native riparian species of vegetation. The native trees and shrubs grow rapidly. Trees will grow 2–3 m. annually, and shrubs will mature and fruit the first year under appropriate conditions. Careful planning will ensure almost immediate use of the area by a large and diverse avian population during most seasons. Plant species used in revegetation efforts should include honey mesquite, cottonwood, willow trees, quail bush, and inkweed. Clearing should be done selectively so that all native trees and all dead trees or trees with large dead snags are left intact. This will make the area attractive to bird species which use snags as perches and cavities for nesting.
Since clearing salt cedar from an area has little deleterious impact on avian communities, we encourage it, provided that at least 25% of the cleared salt cedar is replaced with native vegetation. We must emphasize, however, that native vegetation cannot be expected to thrive by simply planting it anywhere. Autecological conditions such as soil layering and soil density, depth from the surface to the water table, and salinity conditions should be carefully assessed first. Site preparation requirements, such as leveling, extent of tillage, and irrigation system design, must be carefully considered.
Revegetation can be implemented to restore native plant species and enhance wildlife. Water salvage through reduction of evapotranspirative losses and increased streambed capacity to carry floodwater, as a result of overall reduction in the total vegetation, are potential ancillary benefits of revegetation efforts.
Acknowledgments
We wish to thank Marcelett Ector and Cindy D. Zisner for typing the various drafts of the manuscript. Susan M. Cook, Jane R. Durham, Cindy D. Zisner, and Chuck Hunter reviewed the manuscript and provided editorial suggestions. We are grateful to the many field biologists who assisted in collecting field data. The project was jointly funded by the USDI Bureau of Reclamation and the USDI Fish and Wildlife Service through Contract Number 7-07-30-V0009.
Literature Cited
Anderson, B.W., R.W. Engel-Wilson, D. Wells, and R.D. Ohmart. 1977. Ecological study of southwestern riparian habitats: techniques and data applicability. p. 146–155. In : R.R. Johnson and D.A. Jones (ed.). Importance, preservation and management of riparian habitat: a symposium. [Tucson, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43. 217 p. Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo.
Anderson, B.W., and R.D. Ohmart. 1976. Vegetation type maps of the lower Colorado River from Davis Dam to the southerly International Boundary. USDI Bureau of Reclamation, Lower Colorado Region. 27 p. Boulder City, Nev.
Anderson, B.W., and R.D. Ohmart. 1977. Climatological and physical characteristics affecting avian population estimates in Southwestern riparian communities using transect counts. p. 193–200. In : R.R. Johnson and D.A. Jones (ed.). Importance, preservation and management of riparian habitat: a symposium. [Tucson, Ariz., July 9, 1977]. USDA Forest Service GTR-RM-43. 217 p. Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo.
Anderson, B.W., and R.D. Ohmart. 1978. Phainopepla utilization of honey mesquite forests in the Colorado River valley. Condor 80:334–338.
Anderson, B.W., and R.D. Ohmart. 1981. Revegetation efforts along the lower Colorado River. Final Report to the USDI Bureau of Reclamation, in review, Boulder City, Nev.
Anderson, B.W., R.D. Ohmart, and J. Disano. 1978. Revegetating the riparian floodplain for wildlife. p. 318–331. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Georgia, Dec. 11–13, 1978]. USDA Forest Service GTR-WO-12. 410 p. Washington, D.C.
Cohan, D.R., B.W. Anderson, and R.D. Ohmart. 1978. Avian population responses to salt cedar along the lower Colorado River. p. 371–382. In : R.R. Johnson and J.F. McCormick (tech. coord.). Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium. [Callaway Gardens, Georgia, Dec. 11–13, 1978]. USDA Forest Service GTR-WO-12. 410 p. Washington, D.C.
Emlen, J.T. 1971. Population densities of birds derived from transect counts. Auk 88:323–342.
MacArthur, R.H., and J.W. MacArthur. 1961. On bird species diversity. Ecology 42:594–598.