Alder, Cottonwood, and Sycamore Distribution and Regeneration Along the Nacimiento River, California[1]
Allan N. Shanfield[2]
Abstract.—Distribution and regeneration of alder, cottonwood, and sycamore was assessed for much of the Nacimiento River (Monterey County, California). Alder, abundant along narrow reaches, had well-distributed height-classes. Cottonwood, now rare along the river, exhibited low numbers of saplings; overgrazing of cattle is believed primarily responsible for the decline. Sycamore had good regeneration along narrow reaches and poor regeneration on broader bottomlands. It appears unlikely that sycamore status is threatened for it exerts strong clonal regeneration.
Introduction
In recent years the many important values inherent in riparian systems have been recognized (Menke in press; Thomas et al . 1979; Warner 1979; and others). In response, the California Department of Fish and Game (DFG) conducted a riparian vegetation survey of the Nacimiento River, upstream from the Nacimiento Dam (Shanfield 1979). Available literature reveals that little is known of this region, outside of the inventory by Gerdes (1976). This paper draws from Shanfield (1979) and assesses the distribution and regenerative status of alder (Alnus rhombifolia ), cottonwood (Populusfremontii ), and sycamore (Platanusracemosa ). In addition, the effects of cattle browsing upon these species are considered.
The Study Site
The Nacimiento River flows out of the Santa Lucia Mountains in central coastal California. It and the San Antonio River have parallel courses, and each is a major tributary to the north-flowing Salinas River. The study areas are located from the Los Padres National Forest and Fort Hunter Liggett (FHL) borders south to the Monterey County line (fig. 1).
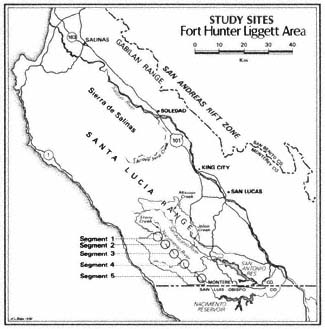
Figure l.
Location of study areas along the Nacimiento River.
Climatically, the area has hot, dry summers and cool, rainy winters. At FHL, the maximum and minimum temperature averages 37.5°C. and 15.8°C. for July, and 15.6°C. and 1.3°C. for January. Annual precipitation has ranged from 58.4 cm. to 73.5 cm. Temperature extremes have ranged from –9.2°C. to 46.8°C. (ibid .).
Much of FHL is periodically disturbed by US Army used each year to reduce fire hazard. An estimated 50% of areas surveyed show evidence of
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, Sept. 17–19, 1981].
[2] Allan N. Shanfield was formerly with the California Department of Fish and Game, Wildlife Management Branch, Monterey, Calif; his present address is in Los Angeles, Calif.
fire effects. Also, cattle grazing is a significant activity that has been modifying the landscape since the early 1800s. In recent years, including the 1976–1977 drought, cattle concentrations were excessively high and the range condition deteriorated. However, in recognition of riparian vegetation as a sensitive resource, the Directorate of Facilities Engineers at the Fort Ord Complex instituted a seasonal grazing program in lieu of a continuous grazing regime, concomitantly lowering animal unit months (AUMs).[3]
Other disturbances include ground squirrels, whose populations had increased because of overgrazing, and an infectious fungus, Anthracnose. This plant pathogen has been reported to hinder sycamore regeneration to an unknown degree (discussed further in Shanfield ibid .).
The Nacimiento River's riparian vegetation typically contains sycamore, willow (Salix spp.), and mulefat (Baccharisviminea ) stands with grasses growing on broader floodplains. In canyons, there are higher densities of alder, willow, and mulefat with an understory of wild rose (Rosacalifornica ), snowberry (Symphoricarpos spp.), poison oak (Rhusdiversiloba ), and herbaceous vegetation. The vegetation of the uplands bordering the riparian zone is typically a mosaic of blue oak (Quercus douglasii )/digger pine (Pinussabiniana ) woodland and oak-savannah grassland interdigitating with chaparral stands. Upstream of the FHL border, the oak-madrone (Arbutusmenziesii ) association becomes important, and bigleaf maple (Acermacrophyllum ) occurs close to the water as the riparian zone assumes a more highly mesic character.
Methods
The riparian zone defined in this study includes areas from the late-spring waterline outward to the furthest mesic riparian species (e.g., willow, sycamore). Five representative study segments, 3.2 km. (2.0 mi.) long, were selected for study in spring 1979. Black-and-white aerial photographs with transparent overlays[4] were used to type-map the vegetation. The ground survey was conducted during spring and summer of 1979.
Sampling units, subsegments having varying lengths, and site selection were predicated upon field-identifiable landmarks (e.g., bend in river, roadways). Within most subsegments the following information was recorded: 1) plant cover as estimated by Daubenmire's (1968) cover-classes utilizing midpoints; 2) census of relative ages of individual plants; and 3) other observations and photo-documented information.
Censused trees were categorized into height-classes as: seedlings (<1.5 m.); saplings (1.6–9 m.), poles (10–17 m.), or mature trees (>17 m.). Cottonwood seedlings were not counted as they were ubiquitous, wherever they did occur, on freshly exposed alluvium and within the river channel. Subsequent to sampling, younger-aged cottonwood and sycamore trees were censused, and trunk cores were taken to determine logical dbh-height bounds for each class. Reported bounds (Stone 1971) for cottonwood ages were used here. A more detailed description of the methods can be found in Shanfield (1979).
Results and Discussion
Census results for size/age groups are presented in table 1. Before any interpretations are made regarding these data, it is appropriate to consider that two populations were present: 1) the population regenerating by clonal spread, wherein stand size/age structures of aboveground stems are different than belowground parts; and 2) the population regenerating from seed (Harper 1977). What appeared to be genetically individual plants (genets) were counted. However, such attempts yield little information regarding the true multiple age-structures of plants having clonal tendencies. This problem is especially inherent in riparian plants. Sycamore and alder exhibited considerable cloning, where an estimated 60–70% of mature and pole-sized trees had either root or trunk sprouts. All censused cottonwoods were individual trees showing slight to zero sprouting, suggesting that cottonwood regeneration is predominantly by seed along the Nacimiento River. This contrasts with Irvine and West's (1979) data showing that cloning is the predominant means of cottonwood regeneration along the Escalante River, Utah.
Species with low levels of regeneration have either: 1) naturally poor regeneration creating an uncertainty as to the "equilibrium" situation; or 2) been subjected to an increase in relative mortality in recent years; or both (Harper 1977; Johnson etal . 1976). Any interpretations, therefore, regarding the regenerative status of the study species are tempered by the fact that only the aboveground populations were sampled. However, both species exhibited a wide range of stem sizes.
Harper (1977) suggested that investigators study and characterize species' ecologies rather than perform statistics on broad age-class averages. Furthermore, an historic approach is necessary to determine riparian vegetation change, and Hastings and Turner's (1965) investigation serves as an excellent model.
Alder Distribution and Regeneration
Alder was most abundant along narrow reaches of study segments 2 and 5. These sites were
[3] Langford, J. 1981. Range Conservationist, Division of Facilities Engineers, Fort Hunter Liggett, Jolan, Calif. Personal communication.
[4] Aerial photographs available at USDA Soil Conservation Service, Salinas, Calif.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
characterized by year-round water, low browsing pressure, and 75–95% cover of alder (fig. 2). Alder was rare along broader floodplains and usually had less than five percent cover. Here, it was usually restricted to east-facing, more mesic streambanks.
Table 1 shows that alder had well-distributed height-classes for all areas except the northern reach of Segment 2. Outside of censused areas, alder had good regeneration both clonally and from seed.

Figure 2.
Typical alder stand along Nacimiento
River, Study Segment 2, looking north.
Cottonwood Distribution and Regeneration
Cottonwood was rare along the Nacimiento River and had less than five percent cover in all subsegments. Trees were distributed close to the water and on higher terraces. Seedlings were ubiquitous, especially on freshly exposed alluvium and in the river bed in summer months.
Most cottonwoods were in older age-classes, and very few saplings were present (table 1). Trees tended towards older ages along wider floodplains (such as Segment 1), while younger age-classes were more common along narrower reaches (Segments 2 and 5). One site of abundant regeneration in Segment 2 (uncensused) had 15–20 saplings on a bank approximately 15 m. long.
Areas of good regeneration typically had the following site characteristics for cottonwood (as well as sycamore) regeneration: 1) narrow riparian corridors (canyons); and 2) low grazing pressure based upon hoofprints, scat evidence, and reduced high-lining, presence of herbaceous vegetation at the water's edge, and ownership information. Glinski (1977) reported excellent regeneration by seed (586 saplings per 0.8 km. of stream edge) along ungrazed reaches in his study areas. Irvine and West (1979) reported that cottonwood was limited to broad flood terraces and regenerated almost exclusively by suckering.
Sycamore Distribution and Regeneration
Sycamore occupied sites closest to the water as well as on outer terraces. It was common on all reaches; its cover values ranged from 19–32% along the riparian zone. Seedlings were present here, contrasting with Griffin's[5] observation that seedlings are absent on Hastings Reserve, which is approximately 55 km. north. Similarly, Glinski (1977) reported an absence of seedlings and only one sapling along 14.4 km. of Sonoita Creek, Arizona, which included both grazed and ungrazed reaches.
Most sycamore stands tended toward older ages in segments 1, 3 (fig. 3), and part of 4 (fig. 4). Narrower, canyon-like sites, such as in segments 2, part of 4, and 5, had younger age-classes. Two sites of active regeneration were found in segments 2 and 5 (fig. 5); these areas had site characteristics similar to those for excellent cottonwood regeneration.

Figure 3.
Mature sycamore and cottonwood
trees along river. Location: segment 1.

Figure 4.
Mature sycamore stands in wash.
Location: segment 4, looking south.

Figure 5.
Typical site of sycamore regeneration in
segment 2. Note bunchgrasses at water's edge.
Effects of Cattle Grazing
Direct Effects
Overgrazing by cattle has probably limited cottonwood sapling establishment along much of the Nacimiento River. Sources elsewhere (Glinski 1977; Reed 1979) indicate that cottonwood is a preferred food, and Glinski has provided strong correlations between sapling establishment and levels of grazing pressure. Locally, the sapling (3- to 15-year) class has been most affected, a timeframe synchronous with periods of heavy grazing and drought along the Nacimiento River. Saplings appeared to be locally restricted to areas of slight to zero cattle impact, such as drainage ditches along the Jolon Road, Salinas River at King City; study sites along segments 2 and 5; and within Mission Creek.
The Mission Creek site (fig. 6 and 7) provides a fenceline comparison that illustrates the degree of vegetation modification attributed to cattle grazing. Similar effects have been noted in the inter-mountain region (Keller etal . 1979; Storch 1979).
Upstream from the fenceline along Mission Creek, where cattle had been excluded, were countless cottonwood (as well as mulefat and willow) saplings which colonized an alluvium bed deposited about 1975 as a result of a dam break. Here, alder and sycamore regeneration was minimal, suggesting that these species occupy separate niches or are very poor interspecific competitors. This stand also suggests that cottonwood has strong potential for sapling establishment when proper seedbed conditions exist in conjunction with reduced grazing pressure.
[5] Griffin, J.R. 1981. Curator and Plant Ecologist, Hastings Reservation, University of California, Carmel Valley.

Figure 6.
View north, ungrazed and fenced portion of Mission
Creek. Rocks at lower left serve as landmark.

Figure 7.
View looking south of grazed portion of Mission
Creek. Rocks in foreground are the same as in
figure 5. Cattle have not been excluded,
and regeneration is greatly reduced.
If grazing were the primary factor limiting tree establishment, then there should be a consistent increase in cottonwood and sycamore regeneration on a larger scale in areas with reduced grazing pressure (segments 2 and 5), but that is not the case. Cottonwood is naturally rare in narrower reaches, along both the Nacimiento and Escalante rivers (Irvine and West 1979), for where there is channelization, there is increased inundation and mechanical damage from water and sediment, fluvial erosion and deposition. Such fluctuations may wash out or bury young plants (ibid .).
Sycamore, in contrast, appears less affected by direct grazing pressure. It is reported to be an unpalatable browse species (Glinski 1977; Martin 1979). Sycamore saplings were absent in areas of reduced grazing pressure (e.g., Hastings Reserve, Mission Creek and those reaches studied by Glinski). It may be that the regenerative success of sycamore is largely independent from grazing pressure. It seems reasonable that other factors may contribute to poor sapling establishment; or sycamore may normally regenerate only by cloning, and once lost from an area may not reestablish itself.
Indirect Effects
Cottonwood and sycamore regeneration and the presence of understory species were typically limited to the water's edge along wide, braided floodplains. Segment 1, for example, had predominantly older, possibly even-aged trees widely dispersed on benches (fig. 3).[6] Much of this area has been burned frequently, and grazing levels were high, as exemplified by the degree of highlining on trees. The soils are reported to be nutrient-poor, and bedrock is often exposed.[6]
Glinski (1977) and Irvine and West (1979) observed that trees rarely occurred on higher ground in their study areas, and it is believed that poor lateral water flows, in conjunction with other factors, have contributed to a "desertification" of riparian terraces. It appears reasonable that seedling establishment in such locales is a tenuous possibility, for young plants must quickly extend their root systems to survive desiccation, fires, and cattle grazing.
In mountain meadow situations, Platts (1981) found that cattle grazing initiated multiple effects upon riparian soils; where livestock trample and compact the soil, the range favors species that exist in areas of lowered water tables. As soils compact and groundcover diminishes, infiltration of water into deep soils becomes poor, surface runoff is increased, and rich topsoil is lost by water and wind erosion.
Much of the Nacimiento River had poor instream cover by comparison to the fenced Mission Creek site (figs. 6 and 7). Where there was less impoundment due to less overhanging vegetation and debris along the stream, there would be reduced lateral flows to deposit nutrient-rich alluvium and recharge water capacities on higher terraces.
Conclusions and Recommendations
In the study area, alder had good regeneration as exemplified by high relative numbers of
[6] Summers, W.J. 1981. Range Conservationist, Natural Resources Management Branch, Naval Facilities Engineers, San Bruno, Calif. Personal conversation.
saplings. Its multiple-stemmed, clonal nature and relative lack of grazing use should ensure its existence.
Cottonwood may be declining along the Nacimiento River because of low recruitment levels. Cattle grazing has probably limited its regeneration and distribution on potentially suitable sites, both directly and indirectly.
Sycamore had various levels of regeneration which are correlated with specific site conditions. Cattle grazing may have indirectly affected sycamore distribution and regeneration along broader bottomlands; however, these data concur with Glinski (1977). Sycamore status will be unthreatened because it can exert strong clonal regeneration, that is, a large percentage of mature trees have root and trunk sprouts that should ensure its survival.
Large-scale regeneration of cottonwood and sycamore along the Nacimiento River could occur as pulses resulting from massive flooding and deposition cycles and coupled with a cessation of limiting factors (e.g., disease, grazing, etc.). This sequence is comparable to Harper's (1977) contention that regeneration occurs in waves rather than as a continuous process.
It is suggested that, where feasible, research and demonstration areas be created along the Nacimiento River where cattle are excluded (Platts 1981). This could provide a control situation where researchers could isolate the many habitat factors affecting riparian vegetation ecology (see Westman [1981] for list of habitat factors). Some suggested areas of research are: 1) what is the recovery time for riparian areas from cattle grazing, and what successional changes occur (see Platts [1981] for discussion of needed riparian/grazing research); 2) what are the effects of fire upon riparian vegetation; 3) how can "normal" regeneration of the studied species be characterized; 4) what is the autecology of the study species; 5) how much outbreeding does sycamore exhibit, and what levels are necessary to ensure its stability over time; and 6) what grazing strategies are most compatible with riparian resources in the California Coast Ranges.
Acknowledgments
I express my gratitude to Drs. Menke and Bartolome for their thoughtful and incisive comments. Also, special thanks to Bruce Elliott for permission to use these data, and to Will Summers, Tim Brothers, and Kris Preston and John O'Leary for their generous input into this paper.
Literature Cited
Daubenmire, R.F. 1968. Plant communities: a textbook of plant synecology. 300 p. Harper and Row Co., New York, N.Y.
Gerdes, G. 1976. A study of the Salinas River and its selected tributaries, California. Unpublished report on file with Department of Fish and Game, Wildlife Management Branch, Monterey, California.
Glinski, R.L. 1977. Regeneration and distribution of sycamore and cottonwood trees along Sonoita Creek, Santa Cruz County, Arizona. p. 116–123. In : R.R. Johnson and D.A. Jones (tech. coord.). Importance, preservation and management of riparian habitats: a symposium. [Tuscon, Arizona, July 9, 1977.] USDA Forest Service GTR-RM-43, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. 217 p.
Harper, J.L. 1977. Population biology of plants. 829 p. Academic Press, New York, N.Y.
Hastings, J.R., and R.M. Turner. 1965. The changing mile. 317 p. University of Arizona Press, Tucson, Arizona.
Irvine, J.R. and N.E. West. 1979. Riparian tree species distribution and succession along the lower Escalante River, Utah. South-western Naturalist 24(2):331–346.
Johnson, W.C. etal . 1976. Forest overstory vegetation and environment on the Missouri River floodplain in North Dakota. Ecol. Monog. 46(1):59–84.
Keller, C.C., L. Anderson, and P. Tappel. 1979. Fish habitat changes in Summit Creek, Idaho, after fencing the riparian area. p. 46–52. In O.B. Cope (ed.). Proceedings of the forum on grazing and riparian/stream ecosystems. 94 p. Trout Unlimited, Inc., Washington, D.C.
Martin, S.C. 1979. Evaluating the impacts of cattle grazing on riparian habitats in the national forests of Arizona and New Mexico. p. 35–38. In : O.B. Cope (ed.). Proceedings of the forum on grazing and riparian/stream ecosystems. 94 p. Trout Unlimited, Inc., Washington, D.C.
Menke, J.W. (ed.). in press. Symposium on livestock, wildlife, fisheries relationships in the Great Basin. [Sparks, Nevada, May, 1977.] USDA Forest Service Pacific South-west Forest and Range Experiment Station, Berkeley, Calif.
Platts, W.S. 1981. Influence of forest and rangeland management on anadramous fish habitat in western North America—effects of livestock grazing. Intermountain Forest and Range Experiment Station, Boise, Idaho.
Reed, J. 1979. Resources-use, abuse, and management. p. 64. In : O.B. Cope (ed.). Proceedings of the forum on grazing and riparian/stream ecosystems. 94 p. Trout Unlimited, Inc., Washington, D.C.
Shanfield, A.N. 1979. Nacimiento River study. District Administrative Report 2. Unpublished report on file with Department of Fish and Game, Wildlife Management Branch, Monterey, California.
Stone, E.C. 1971. The dynamics of vegetation change. Unpublished report prepared for Cal-American Water Company. May be obtained through Department of Forestry and Resource Management, University of California, Berkeley.
Storch, R.L. 1979. Livestock/streamside management programs in eastern Oregon. p. 56–60. In : O.B. Cope (ed.). Proceedings of the forum on grazing and riparian/stream ecosystems. 94 p. Trout Unlimited, Inc., Washington, D.C.
Thomas, J. W., C. Maser, and J.E. Rodiek. 1979. Riparian zones in managed rangelands - their importance to wildlife. p. 21–30. In : O.B. Cope (ed.). Proceedings of the forum on grazing and riparian/stream ecosystems. 94 p. Trout Unlimited, Inc., Washington, D.C.
Warner, R.E. 1979. The California riparian study program phase I: background study and program design for phase II. 177 p. California Department of Fish and Game, Sacramento, Calif.
Westman, W.E. 1981. Factors influencing the distribution of species of Californian coastal sage scrub. Ecology 62(2):439–455.