The Use of Riparian Systems by Aquatic Insects[1]
Nancy A. Erman[2]
Abstract.—Nearly all aquatic insects spend some portion of their lives in riparian zones. Many examples are given of terrestrial stages and activities of aquatic insects. The examples are divided into the areas of feeding, pupation, emergence and mating, and egg laying. Special emphasis is given to recent studies on caddisflies (Trichoptera) in riparian systems of the Sierra Nevada. The examples illustrate the point that knowledge of life cycles is essential for predicting the effect of disturbance in riparian areas on populations of aquatic insects.
Introduction
Most aquatic insects are either directly or indirectly dependent on riparian vegetation at some stage in their life cycles. This dependence has two aspects. The first is that the riparian system affects the aquatic system. The second is that nearly all aquatic insects are terrestrial at some stage in their life cycle and many at several stages in their life cycle (table 1). These terrestrial stages are usually spent close to the water in the riparian zone. It is the second aspect of riparian use that will be covered in this paper, but a brief review of the first is relevant to the theme of this symposium.
Riparian systems act on streams to produce shade and maintain cool summer stream temperature, to provide leaf fall, and to prevent soil erosion (Ross 1963). The smaller the stream, the greater is the effect of the riparian system on it and the more restricted to terrestrial biomes are the aquatic insects (Ross ibid .). In small streams in upper watersheds, terrestrial leaf litter is an important part of the diet of aquatic insects (Cummins 1973). Indirectly, insects farther downstream depend on the same food source when they eat fine particulate organic matter chewed and egested by those upstream. Carnivorous insects are indirectly dependent on riparian vegetation because their prey feed on it. Insects feed also on food produced within the aquatic system—algae and aquatic macrophytes (Merritt and Cummins 1978). The relative importance of allochthonous and autochthonous food sources continues to be discussed (Kaushik and Hynes 1971; Hynes 1975; Minshall 1978; Vannote etal . 1980), but there is no doubt that the riparian zone contri-
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Nancy A. Erman is Staff Research Associate in freshwater ecology, Department of Forestry and Resource Management, University of California, Berkeley, Calif.
butes a large part of the food base to any stream system.
Detailed studies of aquatic insect food habits have shown that many insects are omnivorous and that food needs change with instars (Chapman and Demory 1963; Winterbourn 1971; Mecom 1972; Anderson and Cummins 1979; Erman 1981). Therefore, an insect that may exist on algae produced within the stream in the early instars, may later shred decaying leaves from the riparian zone and later still become carnivorous.
The second aspect of riparian use by aquatic insects, that of their terrestrial stages, can be divided into four areas—feeding and, in the Trichoptera, case building; pupation; emergence and mating; and egg laying. Table 1 is a general representation of life stages of aquatic insects. Many exceptions have been omitted for simplification. It is particularly difficult to classify an order aquatic or terrestrial in any given stage when some orders (Hemiptera, Lepidoptera, Coleoptera, Diptera) have so many semiaquatic members. These members should be kept in mind, because although they are not covered in this paper, they are undoubtedly important in riparian zones. Special emphasis is given here to recent studies on caddisflies (Trichoptera) in riparian systems of the Sierra Nevada.
Feeding on Streamside Vegetation
Most feeding by aquatic insects occurs in the nymphal or larval stage and, as discussed above, in the water. However, one caddisfly, Desmonabethula (Trichoptera: Limnephilidae) leaves the water to feed on several species of semiaquatic plants (Erman 1981). The life cycle is shown in figure 1. The larvae live in slow, unshaded stretches of small spring-streams in the Sierra Nevada and were studied at the Sagehen Creek Field Station (Nevada County). In most instances the larvae are in sections of springstreams associated with fens or minerotrophic peatlands, described elsewhere (Erman and Erman 1975). Larvae leave the water at night to feed on Carex nebraskensis , C . praegracilis , C . rostrata , Calamagrostiscanadensis (leaves and developing flowers), Veronicaamericana , Saxifragaoregana , Dodecatheonalpinum (developing fruits only), and Polygonumbistortoides . In all but the species indicated, larvae strip the tenderest tips and edges of the leaves (fig. 2). Such feeding is typical of many herbivores, probably because of richer nutrient concentrations in those portions of the plants.
Population estimates of Desmonabethula revealed that 14–16% of the population migrated on any one night but that a different segment migrated the following night. All or nearly all of the population may depend on terrestrial feeding at a critical stage in its life cycle Fifth instar larvae had high densities, 1220/m2 and 1662/m2 in optimum stream sections. It may be that D . bethula reaches high numbers
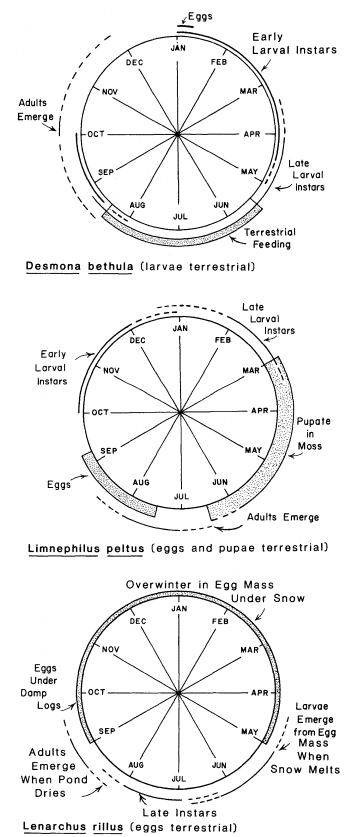
Figure l.
Life cycle of three Trichoptera (Limnephilidae)
in the Sagehen Creek basin (Nevada County,
Calif.) showing aquatic and terrestrial periods.

Figure 2.
The caddisfly larva, Desmona bethula (Trichoptera :
Limnephilidae) feeding at night on emergent
vegetation (Carex spp.). Photo by Jerry Morse.
because it can leave the stream and use a food source unavailable to other aquatic insects.
Another group of caddisfly larvae, Cryptochia spp. (Limnephilidae) has been found on wet wood and leaves at the water's edge as well as in the water (Wiggins 1977). Psychoglypha spp. (Limnephilidae) have been observed at the edge of spring-streams in the Sagehen Creek basin, Sierra Nevada, feeding on Veronicaamericana (N.A. Erman unpublished).
Some species of Lepidostoma (Trichoptera: Lepidostomatidae) are found only along the very edges of streams in the roots of sedges and other aquatic plants. These species are rarely if ever collected in open water. They use root material in their cases and may feed on it (N.A. Erman unpublished). Also Onocosmoecus spp. (Trichoptera:Limnephilidae) live along stream edges and feed on decaying grass that bends into the stream (Wiggins 1977).
There are other caddisflies, particularly in the family Limnephilidae, that live in springs, seeps, temporary streams, and along the edges of larger streams in California, and probably use the riparian zone as a food source. The Limnephilidae is a highly diverse family, important in California because it is dominant at higher elevations in the Nearctic region. The above are only given as examples and should not be considered an inclusive list.
Many of the aquatic and semiaquatic Lepidoptera larvae feed on cattail (Typha spp.), bulrush (Scirpus spp.) or other vascular hydrophytes (Lange 1978). Also some of the aquatic Diptera larvae and some of the aquatic Coleoptera feed on living vascular plant tissue (Doyen and Ulrich 1978; Merritt and Schlinger 1978) though usually the plants are hydrophytes and might not be strictly riparian.
Adult stoneflies (Plecoptera) live on streamside vegetation where species of several families (Taeniopterygidae, Nemouridae, Capniidae, Chloroperlidae, some Perlodidae) feed on epiphytic algae or young leaves and buds (Harper 1978; Jewett 1963; Brinck 1949).
Pupation Outside the Water
All the Megaloptera and aquatic Neuroptera leave the water to pupate on land, usually very close to the water. The alderflies (Megaloptera: Sialidae) burrow into soil along the bank (Azam and Anderson 1969). The dobsonflies and fishflies (Megaloptera:Corydalidae) pupate either in the soil or in decaying shoreline trees or stumps. And the spongillaflies (Neuroptera: Sisyridae) spin silk cocoons usually located in protected terrestrial sites near shore (Evans 1978).
In the Trichoptera terrestrial pupation is the exception but seems to have evolved as an adaptation to life in intermittent streams. Caddisfly larvae leave the water in the final instar and pupate on land as the water evaporates. Examples are Ironoquiaparvula (Limnephilidae) (Ross 1944; Flint 1958), I . punctatissima (Williams and Williams 1975), and I . lyrata (Mackay unpublished).
Limnephiluspeltus (Limnephilidae) leave spring-streams shortly after snowmelt in the Sagehen Creek basin and burrow into moss along the stream edges in the fens (fig. 1). In dry years when spring flow recedes and moss dries out early, some pupae die befor emerging (N.A. Erman upublished).
Nearly all aquatic Tipulidae (Diptera) leave the water to pupate in nearby soil, moss or litter (Byers 1978), and pupation is terrestrial in almost all species of aquatic Coleoptera (Leech and Chandler 1956; Pennak 1978). Usually they pupate in cells excavated by larvae under stones or logs. The Gyrinidae make mud cells on aquatic vegetation (Doyen and Ulrich 1978).
Adult Emergence and Mating in the Riparian Zone
By the very fact of emerging from the water most aquatic insects "use" the riparian zone, at
least for resting, but in some the use is more specific than in others.
The mayflies (Ephemeroptera) emerge from the water as subimagoes (winged adults but sexually immature). They may perch on streamside vegetation for only a few minutes or for as long as 48 hours (about 24 hours for most species) before molting to the imago (Day 1963; Edmunds etal . 1976).
Adults of the dragonfly genus Aeshna (Odonata:Aeshnidae) tend to hang in the shade on the underside of leaves of trees in hot weather (Smith and Pritchard 1963).
Males of some Plecoptera wait on the shoreline for females to emerge. They drum on dry leaves, rocks, and moss by striking their abdomen on the substrate apparently to attract females. Mating takes place on the ground (Brinck 1949).
Adult caddisflies, Desmonabethula , fly up into lodgepole pines (Pinuscontorta ) along streams and peatlands immediately upon emerging from the water (Erman 1981). This same behavior has been observed in Heteroplectron californica (Trichoptera:Calamoceratidae), and Limnephiluspeltus is known to mate on lodgepole pines near wetlands (N.A. Erman unpublished).
Several families of aquatic Diptera fly in mating swarms above vegetation, and most adults of aquatic Diptera use riparian vegetation or ground near the water as resting sites (Merritt and Schlinger 1976).
Egg Laying in the Riparian Zone
Eggs of aquatic insects laid outside the water must have a mechanism for entering the water either as an egg or as a newly hatched larva. Therefore, they must be close to the water, often on overhanging vegetation.
Some species of Odonata attach their eggs onto plant stems, while others deposit them in plant tissue above drying ponds where they do not hatch until the ponds are refilled (Westfall 1978). Certain damselflies use emergent vegetation in another way during egg laying. The male clings to a plant, grasps the female by the prothorax with his terminal abdominal appendages, and lowers her into the water while she deposits eggs (Smith and Pritchard 1963).
All of the Megaloptera and aquatic Neuroptera lay their eggs out of water, often on leaves or branches overhanging the water. Eggs hatch and larvae fall or crawl into the water (Gurney and Parfin 1959; Azam and Anderson 1969).
Several Trichoptera species lay eggs outside the water, and as in the case of terrestrial pupation, it seems often to be another adaptation to drying conditions. Eggs are laid along the edges of drying streams and ponds, often under damp logs. Larvae hatch but remain in the gelatinous egg mass until the stream or pond is reflooded (Novak and Sehnal 1963; Wiggins 1973; Wiggins etal . 1980). An example of this adaptation is known for Lenarchusrillus in the Sierra Nevada (N.A. Erman unpublished), shown in figure 1.
In other Trichoptera species the gelatinous egg masses are laid on stems above the water. When the larvae hatch and become active the gelatinous matrix thins, liquifies, and flows down the stem into the water (Wiggins 1973).
The female of Yphriacalifornica (Trichoptera:Phryganeidae) swims underwater and attaches egg masses to the underside of overhanging moss in small streams in the Sierra Nevada (N.A. Erman unpublished).
It is highly likely that as caddisfly behavior is studied in more specific detail, other species with aquatic/terrestrial adaptations may be revealed because Trichoptera is an order in which several members have reinvaded the land as larvae (Tomaszewski 1973; Mackay and Wiggins 1978; Erman 1981).
Beetles of some of the aquatic and semiaquatic families, i.e. Dytiscidae, Helodidae, Georyssidae, Heteroceridae, Hydraenidae; oviposit in wet, shoreline places (Leech and Chandler 1963; Doyen and Ulrich 1978).
Aquatic Diptera often attach eggs to rocks or aquatic vegetation (Teskey 1978). Some of the Simuliidae oviposit on wet surfaces such as grass trailing in the water (Peterson 1978), and Chironomidae deposit gelatinous egg masses on emergent substrates or vegetation (Coffman 1978). The aquatic Diptera, like the Trichoptera, is an ecologically diverse and understudied group that will no doubt reveal more aquatic/terrestrial interactions when it is further investigated.
Management Implications
It is evident from the foregoing discussion that disturbance in the riparian zone will ultimately affect the large and varied group of aquatic insects present in any reasonably healthy stream or lake. Aquatic insects have intricate life cycles which if interrupted at any point may mean the disappearance or reduction of a given species in a given area. While many insects have life cycles with great flexibility, enabling them to respond to environmental conditions, i.e. diapause which delays development of an egg, larva, pupa, or adult until conditions are favorable, it must be remembered that such flexibility has evolved over long periods in response to gradually changing conditions. Such activities as channeling streams, logging, building dams, grazing livestock, drawing water out of streams, and putting in riprap have the potential for sudden interference with life cycles.
Dams are so disruptive to all stream life that there is no point in discussing them in this context. The effects of logging on stream life are being considered elsewhere (Mahoney and Erman 1981).
An insect that needs soil along a bank for a pupation chamber will not find it in a channelized stream or one with riprapped banks. Larvae that must immediately drop into water upon hatching may be eliminated if vegetation is destroyed because adults will not find suitable egg-laying sites. It is worth noting that Desmonabethula , the first caddisfly larva known to migrate daily from the water to feed, was found in a study area where grazing has been eliminated for many years. Its habitat, springstream in peatlands, is also a favorite of livestock in the Sierra Nevada. It may be no coincidence that these larvae have never been reported from any other area.
The more we know about life cycles, behavior, and feeding habits of individual species of aquatic insects, the more accurate can be our predictions on the effects of changes in the riparian zone. It is already clear that the riparian zone and aquatic insects are connected in an inseparable and intricate web of relationships.
Acknowledgements
I thank Don C. Erman for reviewing the manuscript and Jerry Morse for photographic work.
Literature Cited
Anderson, N.H., and K.W. Cummins. 1979. Influences of diet on the life histories of aquatic insects. J. Fish. Res. Board Can. 36:335–342.
Azam, K.M., and N.H. Anderson. 1969. Life history and habits of Sialisrotunda and S . californica in western Oregon. Ann. Ent. Soc. Am. 62:549–558.
Brinck, P. 1949. Studies on Swedish stoneflies. Opusc. Ent. Suppl. 11:1–250.
Byers, G.W. 1978. Tipulidae. p. 285–310. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Chapman, D.W., and R.L. Demory. 1963. Seasonal changes in the food ingested by aquatic insect larvae and nymphs in two Oregon streams. Ecology 44:140–146.
Coffman, W.P. 1978. Chironomidae. p. 345–376. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Cummins, K.W. 1973. Trophic relations of aquatic insects. Ann. Rev. Ent. 18:183–206.
Day, W.C. 1963. Ephemeroptera. p. 79–105. In : R.L. Usinger (ed.). Aquatic insects of California. 508 p. University of California Press, Berkeley, Calif.
Doyen, J.T., and G. Ulrich. 1978. Aquatic Coleoptera. p. 203–231. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Edmunds, G.F., Jr., S.L. Jensen, and L. Berner. 1976. The mayflies of North and Central America. 330 p. University of Minnesota Press, Minn.
Erman, D.C., and N.A. Erman. 1975. Macroinvertebrate composition and production in some Sierra Nevada minerotrophic peatlands. Ecology 56:591–603.
Erman, N.A. 1981. Terrestrial feeding migration and life history of the stream-dwelling caddisfly, Desmonabethula (Trichoptera: Limnephilidae). Can. J. Zool. 59:1658–1665.
Evans, E.D. 1978. Megaloptera and aquatic Neuroptera. p. 133–145. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Flint, O.S. 1958. The larva and terrestrial pupa of Ironoquiaparvula (Trichoptera: Limnephilidae). J. New York Entomol. Soc. 66:59–62.
Gurney, A.B., and S. Parfin. 1959. Neuroptera. p. 973–980. In : W.T. Edmondson (ed.). Freshwater biology (2nd ed.). 1248 p. Wiley, New York.
Harper, P.P. 1978. Plecoptera. p. 105–118. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Hynes, H.B.N. 1975. The stream and its valley. Verh. Internat. Verein. Limnol. 19:1–15.
Jewett, S.G., Jr. 1963. Plecoptera. p. 155–181. In : R.L. Usinger (ed.). Aquatic insects of California. 508 p. University of California Press, Berkeley, Calif.
Kaushik, N.K., and H.B.N. Hynes. 1971. The fate of dead leaves that fall into streams. Arch. Hydrobiol. 68:465–515.
Lange, W.H. 1978. Aquatic and semiaquatic Lepidoptera. p. 187–201. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Leech, H.B., and H.P. Chandler. 1963. Aquatic Coleoptera. p. 293–371. In : R.L. Usinger (ed.). Aquatic insects of California. 508 p. University of California Press, Berkeley, Calif.
Mackay, R.J., and G.B. Wiggins. 1979. Ecological diversity in Trichoptera. Ann. Rev. Entomol. 24:185–208.
Mahoney, D., and D.C. Erman. 1983. The role of streamside bufferstrips in the ecology of aquatic insects. In : R.E. Warner and K.M. Hendrix (ed.). [University of California, Davis, September 17–19, 1981]. University of California Press, Berkeley.
Mecom, J.O. 1972. Feeding habits of Trichoptera in a mountain stream. Oikos 23:401–407.
Merritt, R.W., and K.W. Cummins. 1978. An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Merritt, R.W., and E.I. Schlinger. 1978. Aquatic Diptera. Part two: adults of aquatic Diptera. p. 259–283. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Minshall, G.W. 1978. Autotrophy in stream ecosystems. BioScience 28:767–771.
Novak, K., and F. Sehnal. 1963. The development cycle of some species of the genus Limnephilus (Trichoptera). Cas. Csl. Spol. Ent. 60:68–80.
Pennak, R.W. 1978. Freshwater invertebrates of the United States, second edition. 803 p. John Wiley & Sons, New York.
Peterson, B.V. 1978. Simuliidae. p. 331–344. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Ross, H.H. 1944. The caddis flies, or Trichoptera, of Illinois. Bull. Ill. Nat. Hist. Survey 23(1):1–326.
Ross, H.H. 1963. Stream communities and terrestrial biomes. Arch. Hydrobiol. 59:235–242.
Smith, R.F., and A.E. Pritchard. 1963. Odonata. p. 106–153. In : R.L. Usinger (ed.). Aquatic insects of California. 508 p. University of California Press, Berkeley, Calif.
Teskey, H.J. 1978. Aquatic Diptera. Part one: larvae of aquatic Diptera. p. 245–257. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Tomaszewski, C. 1973. Studies on the adaptive evolution of the larvae of Trichoptera. Acta Zool. Cracoviensia 18:311–398.
Vannote, R.L., G.W. Minshall, K.W. Cummins, J.R. Sedell, and C.E. Cushing. 1980. The river continuum concept. Can. J. Fish. Aquat. Sci. 37:130–137.
Westfall, M.J., Jr. 1978. Odonata. p. 81–98. In : R.W. Merritt and K.W. Cummins (ed.). An introduction to the aquatic insects of North America. 441 p. Kendall/Hunt Publ. Co., Dubuque, Iowa.
Wiggins, G.B. 1973. A contribution to the biology of caddisflies (Trichoptera) in temporary pools. Life Sci. Contr., Roy. Ont. Mus. 88:1–28.
Wiggins, G.B. 1977. Larvae of the North American caddisfly genera (Trichoptera). 401 p. University of Toronto Press, Toronto.
Wiggins, G.B., R.J. Mackay, and I.M. Smith. 1980. Evolutionary and ecological strategies of animals in annual temporary pools. Archiv. fur Hydrobiologie Suppl. 58:97–206.
Williams, D.D., and N.E. Williams. 1975. A contribution to the biology of Ironoquia punctatissima (Trichoptera:Limnephilidae). Can. Ent. 107:829–832.
Winterbourn, M.J. 1971. An ecological study of Banksoilacrotchi Banks (Trichoptera: Phryganeidae) in Marion Lake, British Columbia. Can. J. Zool. 49:637–645.