1
Shoreline And Other
Naturally Open
Habitats
Mangrove Swamps
By definition, mangroves are trees rooted in substrates that are flooded by seawater, either constantly or periodically. Mangroves belong to many different angiosperm families, some of which also include upland genera, but all the mangrove genera are tightly confined to their saline swamp habitat. Most mangrove genera are tropical, with a minority ranging into the subtropics. Members of the genus Avicennia, a tree belonging to the Verbena family, range farthest into temperate regions. Within their latitudinal limits, mangroves have very broad edaphic and climatic tolerances. Because they are able to draw water molecules and nutrients from the sea, they are remarkably indifferent to substrate, growing in rock crevices and many kinds of sand and mud. Where there is good circulation of seawater, they are also remarkably indifferent to rainfall, growing adjacent to rain forests, deserts, and everything between. Nevertheless, suitable mangrove habitats are extremely discontinuous because of the need of shelter from wave action for seedling establishment.
Within a mangrove forest, species patterns often show striking zonation, which has often been interpreted to be the result of autogenic succession: a pioneer fringe advances seaward, traps sediment, builds up land, and prepares the way for advance of the next zone. However, case histories show the dynamics are usually more complex.
River Deltas, Southern Gulf of Mexico
(Thom 1967; West et al. 1969)
The great uncontrolled rivers of Tabasco and Campeche have provided a dynamic habitat complex in which mangrove distribution patterns are continually changing. In historical time, active river mouths have switched between some distributary channels that fan out over the deltas. During floods, active distributaries build ever higher and wider natural levees along their channels. The sand they pour into the Gulf of Mexico is carried by longshore currents to be laid up by waves as a series of accretion beach ridges parallel to the shore. The constant loading of additional sediment causes regional subsidence of the whole delta. When a distributary is becoming inactive, the old channel is slowly filled with fine sediment deposited by general floods during the winter rains. During the dry season, saline water invades abandoned stream channels and lagoons, and penetration increases during the years as subsidence goes on. Water levels are less affected by the trivial lunar tides than by changes in wind direction, the highest levels coming during coincidence of maximum river discharge and strong north winds during winter. These northerly storms, called nortes, also cause severe beach erosion. After the sand supply has been diverted by a shift in a river mouth, huge areas of beach ridge systems near the abandoned mouth are eventually lost to the sea.
The dominant mangrove species in this region are Rhizophora mangle, Avicennia germinans, and Laguncularia racemosa ; all produce nondormant seedlings capable of long flotation. The mangroves do not colonize natural levees being built by active distributaries; there the first colonists are marsh plants, such as Phragmites, Spartina, and Typna spp. When a distributary is being abandoned and becoming saline, Rhizophora seedlings floating in from the sea usuallv colonize the banks. The tangle of stilt roots of the Rhizophora fringe catches debris and sediment, and the mangroves add organic detritus of their own. As the channel fills and narrows, the Rhizophora fringe widens by establishment of new seedlings, while Avicennia and Laguncularia seedlings join the older Rhizophora in the rear, forming a mixed mangrove forest. As long as peat formation under the mixed mangrove forest compensates for subsidence, the species may persist in situ indefinitely unless the forest is destroyed by retreat of the seashore or is overrun by a new active stream channel.
A different geomorphic—biotic sequence begins on the back side of the natural levees, which slope gradually to mudflats along seasonally saline lagoons. As the mudflats are built up by sedimentation during floods, they are first colonized by Spartina and Batis maritima, a prostrate perennial halophyte. Here Avicennia is usually the pioneer mangrove, with the other
two joining in as sedimentation proceeds. When the distributary becomes inactive and sedimentation is shut off, subsidence will cause the mangrove fringe, with Avicennia in the lead, to move toward the crest of the sinking levee. Lagoons enlarged by subsidence eventually have enough fetch for generation of waves, which undercut the mangroves and wash them away.
Thom (1967) postulated a variety of other geomorphic sequences in this deltaic complex. His general conclusion was that in certain situations, mangroves trap sediment and lay down peat so that vegetationally controlled, autogenic succession may temporarily prevail, but that over the long run habitat changes controlled by the mangroves are overwhelmed by a grand geomorphic cycle controlled by the rivers and the sea.
Coral Cays, Belize
(Stoddart 1962, 1963, 1969)
Mangroves occupy a totally different geomorphic setting on the other side of the Yucatan peninsula, where they grow on coral cays with no river or terrestrial sediments. The cays lie on the barrier reef and on atoll-like structures beyond. The cays most exposed to wind and surf have beach ridges built of coarse coral debris, which are colonized by species discussed in the next chapter. On these exposed cays, mangroves are confined to a fringe on the leeward shore. On more sheltered cays, typically shoals with no dry land, mangroves occupy the whole area. Rhizophora mangle is generally strongly dominant, particularly on the outer margins of the mangrove forest, joined by Avicennia germinans and Conocarpus erectus on slightly higher areas. Laguncularia racemosa occurs infrequently.
Autogenic succession, with mangroves advancing as they trap sediment and lay down peat, cannot be a general process in these cays. Some of the mangrove cays apparently have permanent shorelines with rock foundations rising from fairly deep water. There, mature mangroves grow to the edge with no seedling colonization beyond. Other unconsolidated cays are gradually migrating shoreward; whether they are constant in area, increasing, or disappearing probably depends on the reef flat topography, sand supply, waves, and currents, not on the vegetation.
Catastrophic changes in mangrove patterns were documented by Stoddart's (1962, 1963, 1969) surveys before and after Hurricane Hattie in 1961. His 1960–1961 maps of the cays show vegetation patterns that may have been in approximate equilibrium. Most of the cays had not had a major hurricane strike for 30 years; the southernmost part of the surveyed area may have had hurricane damage 15 years before. The center of the 1961 cyclone passed directly over the cays. Sustained wind speeds were estimated at 250
km/hr with gusts to 320 km/hr. Atmospheric pressure was so low that over an area about 100 km in diameter, the sea rose about 5 m above normal level. Huge waves were superimposed on this storm surge.
The storm broke branches and stripped leaves from mangroves over a wide swath. Stoddart's 1965 resurvey found mangroves were generally dead within a swath about 65 km wide, the only survivors being in very small patches on the leeward sides of the larger cays. Within the dead stands, the ground was still occupied by the old roots and there were a few seedlings. New seedlings were establishing mainly in new bare areas.
Great Barrier Reef, Queensland
(Hopley 1982; Stoddart 1980; Stoddart et al. 1978)
Queensland, Australia, has a much richer complement of mangroves than the neotropics, with 27 species belonging to many different angiosperm families at least locally dominant. All are sea dispersed and shared with the East Indies. With ocean swell shut off by the Great Barrier Reef and a tidal range of 3 to 6 m, mangrove swamps require only minimal topographic shelter and are very extensive.
Within a mangrove stand, species commonly show rather regular zonation, which has evoked deductive models of predictable, autogenic succession. However, where distribution patterns have actually been monitored over time, in some cases by detailed remapping over nearly 50 years, irregular and unpredictable changes were found. Rather than actively advancing by autogenic succession, the mangroves generally passively follow changes in reef morphology with coral growth and destruction.
Naturalized Mangroves, Hawaii
(Wester 1982)
Hawaii is better situated to receive drift from Alaska than from the tropical Pacific. It had no native mangroves, the nondormant seedlings evidently being unable to survive the circuitous voyage up the coast of Asia, across the northern Pacific, and back down to the tropics.
In 1902, the American Sugar Company introduced Rhizophora mangle from Florida to Molokai, Hawaii (fig. 1), where it was planted on the south coast in an attempt to stabilize sediment produced by erosion on the slopes of the island. Seedlings were transplanted from Molokai to Oahu and have since volunteered on the islands of Hawaii, Maui, Lanai, and Kauai, evidently
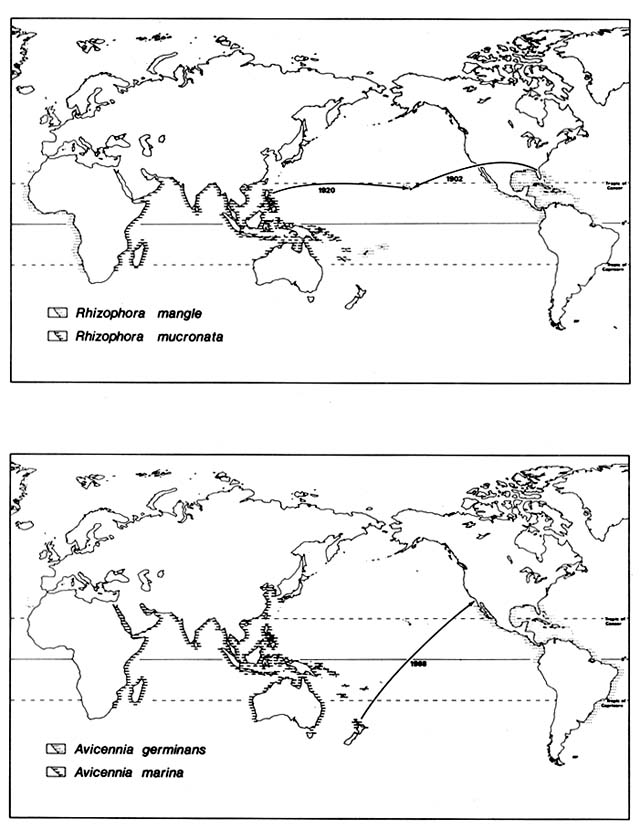
Figure 1. Recent Introductions of Certain Mangroves Outside Their Native Ranges. Mangrove species
generally are well adapted for ocean disposal and have very broad natural ranges. They were, however,
unable to reach some suitable habitats as shown by naturalization following recent artificial introduction.
spreading within the archipelago by natural drift. The neotropical button mangrove, Conocarpus erectus, was also planted in various places along the coast but has not naturalized.
In 1922, four Philippine mangrove species were introduced by the Hawaiian Sugar Planters Association and planted on mudflats around Oahu. Ceriops tagal promptly died; Rhizophora mucronata and Bruguiera parviflora survived for a few years. Bruguiera gymnorhiza has survived and reproduced in several sites on the Oahu coast, in spite of much exploitation of its flowers for leis.
Some of the Hawaiian mangrove colonies depend on artificially modified habitats. The prehistoric rock-walled fish ponds, built in the sea by the Polynesians, offer sheltered habitats for dense mangroves. Mangroves infest canals, including the Ala Wai behind Waikiki, where they must be periodically removed. Extensive stands occupy deltas below sugar mills, where sediment from cane washing is deposited. A 40-acre stand of Rhizophora mangle colonized such a delta in Pearl Harbor between 1951 and 1975. However, other colonies are established on shores naturally sheltered by reefs and embayments. Presumably, these would have been colonized prehistorically if viable seedlings had arrived.
Naturalization of Avicenna, California
(Moran 1980)
Several neotropical mangrove species range naturally up the Pacific coast of Mexico across the Tropic of Cancer. They extend above 29°N latitude in the Gulf of California, but none extend above 27°N on the Pacific side of Baja California.
In 1963, two of these species, Avicennia germinans and Lagunclaria racemosa, were planted experimentally at about 32.5°N latitude in California. The site is a salt marsh dominated by Spartina foliosa on Mission Bay in San Diego. It is administered as a nature reserve by the University of California, San Diego. Neither of these mangroves succeeded, although one Laguncularia survived for several years.
In 1968, seedlings of two Indo-Pacific mangroves were planted in the same marsh: Aegiciras corniculatum from tropical Queensland and Avicennia marina from 37°S latitude in New Zealand. The Aegiciras has survived and flowered but not reproduced. Avicennia marina established thousands of seedlings, now in the third generation; its success threatened the native biota of the marsh. Since 1976, university students have stopped Avicennia reproduction by removal of flowers and seedlings; in 1979 about 12,000 rooted seedlings were pulled up, but a few were left.
Comment
Normal dispersal of mangroves is entirely by ocean currents. Many species are viviparous, that is, seedlings germinate before release from the mother tree and continue to develop while floating. With light, water, and nutrients available, dormancy is pointless. Seedlings occasionally strike root in rock crevices, even on open shores. On loose substrates, they need quiet water. Even where there are only gentle waves, it is common to find dead and dying mangrove seedlings stranded in the drift zone. on the new island of Krakatau (discussed in the section on Fresh Lava and Pyroclastic Deposits), live seedlings of various mangrove species have repeatedly been stranded, but have failed to establish for lack of shelter.
Once established, mangroves are very resistant to storm and wave damage. The catastrophic destruction in the Belize cays (discussed above) is an extreme case. The same hurricane struck Grand Cayman Island less violently than Belize, but Grand Cayman had heavy surf and winds over 100 km/hr; these had little effect on the mangroves. Part of the distribution of mangroves is too close to the equator to be subject to any cyclonic storms. In some sites, mangroves survive in situ indefinitely, as shown by deep peat deposits. However, many populations are only temporary and are inevitably destroyed by gradual or catastrophic habitat changes.
Both the highly disjunct spatial pattern and the temporary survival of local populations imply strong natural selection for dispersibility and perhaps for long-range dispersibility. It is sometimes argued that plants do not evolve adaptations for long-range dispersal through natural selection but that long-range dispersal happens only occasionally and incidentally through adaptation for local dispersal. However, in the case of species obligately dispersed by ocean currents, there is complete elimination of any disseminules that do not complete the voyage to another shoreline. Seeds dropped at random in wind or by bird dispersal have some chance of survival, but seeds that sink at sea do not. Selection in mangroves would not favor long-range dispersal per se but rather long buoyancy, regardless of the rate of drift and distance covered. The more sites, near or far, that a seed or seedling can probe, sometimes by repeated stranding and refloating, the better chance of finding a safe place to take root. In the tropical Indo-Pacific region, which is the center of mangrove diversity, archipelagos of islands offer all degrees of spatial disjunction between suitable sites. Also, the seasonal reversals of monsoonal winds and ocean currents allow genetic feedback from successful distant colonies.
Not all mangroves are known to be capable of long ocean voyages, but various species have ranges spanning the Indian and western Pacific oceans; others span the Atlantic. That their dispersal capacities are not unlimited is shown by the lack of any pantropical species and by the absence of mangroves from Hawaii before the introductions discussed above.
Tropical Sea Beaches
Coral Cays, Belize
(Stoddart 1962, 1963; Stoddart et al. 1982)
Supratidal vegetation, as well as the mangroves discussed above, was mapped on coral cays in Belize just before the devastating hurricane of 1961. The interval since the last storm had probably been long enough for the beach vegetation to reach near equilibrium. The more exposed beaches had a sparse, low outpost vegetation of perennial herbs and vines, all well adapted for long-range ocean dispersal. Many are pantropical, including a beach grass, Sporobolus virginicus ; a beach morningglory, Ipomoea pes-caprae ; and sea purslane, Sesuvium portulacastrum . Others are also widespread but only within the Caribbean region, for example, a spider lily, Hymenocallis littoralis , and a bushy spurge, Euphorbia mesembrianthemifolia .
The larger cays had beach ridge thickets of spray-swept shrubs and trees; these are adapted for both sea and bird dispersal. The dominants included some pantropical species, such as Suriana maritima and Sophora tomentosa , but most are neotropical, such as Thrinax radiata, Coccoloba uvifera, Cordia sebestena, Bursera simaruba, Conocarpus erectus, Tournefortia gnaphalodes, Borrichia arborescens . The East Indian almond, Terminalia catappa , was naturalized on some cays. Many had been largely cleared for coconut plantations.
This beach vegetation was almost identical to that of the Cayman Islands. Before striking Belize, Hurricane Hattie passed close enough to Grand Cayman to cause washing out of the outer few meters of vegetation on some beaches, but the recovery was completed in about 5 years.
On the Belize cays, Hurricane Hattie caused catastrophic damage to beach vegetation over a swath about 65 km wide. Destruction was not as total as in the mangroves. Outpost beach vegetation was damaged mainly by storm waves reworking the substrate. In the most extreme cases, vegetated cays as long as 120 m were completely washed away and have not since reappeared above the sea. More commonly, the outpost vegetation merely lost some ground to beach erosion and washovers or was buried under fresh coral rubble. Parts of the cays were stripped of all loose sand and shingle down to beachrock. In places, losses were selective: waves swept away Ipomoea and Sesuvium but left Sporobolus , and Hymenocallis reemerged where neighboring species were killed by burial.
The beach ridge thickets suffered less from direct wave action than from defoliation and breakage. Here, also, losses were partly selective, Cordia being uprooted more often than neighboring species. The native Thrinax palm stood up to the wind better than the planted Cocos .
Within 6 months, outpost species were recolonizing some of the bare sand, either vegetatively or by seedlings; Sporobolus, Sesuvium , and Ipomoea were among the first. Seedlings of a thicket shrub, Tournefortia , were common. The former distinction between zones was blurred, with outpost species appearing inland of their former position. Some weedy annuals that were not noted on beaches before the hurricane, for example, Portulaca oleracea and Cenchrus incertus , were prominent as early colonizers of fresh sand.
By 4 years after the storm, there was vigorous but patchy colonization of loose sand by both outpost and thicket species. All the outpost species that were common before the storm were well represented and were displacing Portulaca . Among the shrubs that had been prehurricane dominants, Tournefortia was best represented by seedlings; there were also some seedlings of Suriana, Sophora, Thrinax, Conocarpus , and Borrichia . The other thicket species were present as survivors but not as recolonists. Areas of stripped beach rock, coarse shingle, and mats of dead coconut roots remained mostly bare.
By 11 years after the storm, effects of the hurricane were no longer generally obvious except for relic dead and broken trees. Resurveys commonly found more species than the prehurricane roster, perhaps because of more thorough search. However, only a few seedlings of some formerly important thicket species had appeared: Coccoloba, Cordia, Bursera , and the naturalized Terminalia .
Comparative mapping over the same 11-year time span extended to cays south of the swath of Hurricane Hattie. These showed less change, of course, but they still showed quite dynamic cay morphology and vegetation patterns. The maps usually showed only single dominants for each area, so changes may be quantitative rather than advances and retreats of species borders. However, they leave no doubt that micromigrations have gone on, partly as recovery from a storm 15 years before Hattie and partly as a result of chronic instability of sand cays. During the decade, a few species became extinct on some of the smaller cays; some of these had been present originally as individual bushes or trees.
Coral Sand Beaches, Mauritius
(McIntire and Walker 1964; Sauer 1962)
Mauritius is a large, rugged volcanic island, but along with basalt sea cliffs and boulder shores, it has extensive coral sand beaches behind fringing reefs.
Early in 1960, after 15 years without any significant storms, Mauritius was raked by two strong cyclones; the second, Carol, was probably the most
intense in the recorded history of the southwestern Indian Ocean. Instrumentally recorded wind velocities at several stations on Mauritius reached 240 km/hr. Waves superimposed on the storm surge left piles of drift 3 to 4 m above normal wave reach on exposed coasts.
A few months before the storms, the coastal vegetation had been surveyed with belt transects and maps of small sample areas; the vegetation was then in apparent equilibrium and neatly zoned. Outpost vegetation on coral sand beaches was dominated by the same pantropical herbs and vines as on Belize cays. Beach ridge thickets also had the same pantropical shrubs, but two of the dominants were Indo-Pacific species, Tournefortia argentea and Scaevola taccada . Inner beach ridges behind the thickets were generally planted with an introduced timber tree, Casuarina equisetifolia . There were only a few coconut plantations.
Here, as in Belize, destruction by the hurricane of beach outpost vegetation was mostly due to reworking of the sand substrate by storm waves rather than to direct damage to the plants. This was dramatically evident where the same species survived pounding by storm waves where rooted in crevices on basalt but were completely swept away where rooted on sand. Waves also bit into the outer margins of thickets on loose sand and shingle ridges, but survivors of all the species were left in each site.
By contrast, destruction of Casuarina trees extended far beyond the reach of the waves, and mortality in large areas was nearly total. The coconut plantations had mortality of only about 25%.
Three months after Cyclone Carol, a resurvey showed complex changes in beach profiles but little change in shoreline positions because sand had been redeposited as the storm abated. Most of the area reworked by waves was still completely bare, but recolonization was beginning. The commonest new colonists of the outpost zone were Canavalia maritima seedlings and vegetatively propagating Ipomoea pes-caprae and Cyperus stoloniferus , the latter more abundant than before the storm. Some beaches were invaded by seedlings of inland weeds, such as the Mexican prickly poppy, Argemone mexicana , perhaps aided by low salinity after the heavy rains. Except for these weeds, recolonization of the outpost zone was invariably by species that had been present in the same sites before the storm. This outcome was not inevitable, since Ipomoea pes-caprae was the only species that had been present in over 50% of the sites. The storm was evidently ineffective in coastwise dispersal within the outpost zone, probably because seed afloat during the storm was stranded mainly in the thicket zone or farther inland.
In the thicket zone, a few survivors of all the previously present species were found in almost every site; the dominant Tournefortia and Scaevola were resprouting vigorously where damaged. Shrub and tree seedlings were generally few.
Where the storm waves had left bare sand back in Casuarina and Cocos
plantations, there were many new seedlings of species not present before the hurricane. These included all the common outpost and thicket species and also some inland weeds, for example, Portulaca oleracea and Ricinus communis , and crops, for example, Nicotiana tabacum and Ipomoea batatas . Drift stranded during the storm evidently contained a mishmash of propagules derived partly from excavation of beaches and partly from flooding rivers.
During 1961 and 1962 several other cyclones passed near enough to Mauritius to produce heavy surf and beach erosion, mainly on the west coast. The strongest, in 1962, had winds gusting to 200 km/hr. When revisited in 1963, west coast beaches showed additional losses of shrubs that had survived Carol in 1960, but new colonies of outpost herbs, especially Sporobolus and Cyperus , had appeared. On the east coast, recolonization by these and other outpost herbs and vines was further advanced; there were also scattered seedlings of thicket shrubs, mainly Scaevola . The wreckage of the Casuarina plantations had mostly been converted to charcoal, and some replanting had begun.
Pyroclastic Beaches, Krakatau
(Docters van Leeuwen 1936)
After the cataclysmic eruption of Krakatau in 1883, the first expedition to the new island found no sign of plant life. After 15 years, the beaches of volcanic ash and sand had acquired a rich flora including outpost vines, such as Canavalia maritima and Ipomoea pes-caprae ; thicket shrubs, such as Scaevola taccada and Morinda citrifolia ; and beach trees, such as Cocos nucifera, Casuarina equisetifolia, Terminalia catappa, Calophyllum inophyllum , and Pongamia pinnata . Within another 10 years, the island had nearly all the species to be found on beaches of any tropical Indo-Pacific island. The most notable missing species were Tournefortia argentea and Suriana maritima. Tournefortia seedlings have been found occasionally on Krakatau beaches but have not survived. Suriana has not been reported, although there can be little doubt that its sea-dispersed seeds can reach the island. These two species are probably barred from the island by unsuitability of the volcanic substrate; they generally grow on coral and limestone.
The rapid colonization by sea-dispersed beach species is explainable partly by the location in the Sunda Strait between Sumatra and Java. Ocean currents there vary greatly with tides and monsoonal wind shifts, so the island receives drift from both Sumatra and Java, as well as from more distant sources. Success of some species was probably partly due to symbiosis with nitrogen-fixing bacteria; these species include not only the legumes Canavalia and Pongamia but also Casuarina .
Introduced Indo-Pacific Species, Mauritius
(Sauer 1961, 1967b )
Several Indo-Pacific shrubs and trees that are now naturalized along the Mauritius coast would probably be mistaken for natives without the historical record of their introduction.
Mauritius was uninhabited and visited only occasionally by Portuguese and Dutch ships before being temporarily colonized by the Dutch in the seventeenth century. Permanent settlement began with arrival of the French in 1715. Soon after, Mauritius began playing a leading role in the great exchanges of tropical plants between botanical gardens of European colonial powers. Its great botanical gardens at Pamplemousses and Reduit were world famous. Beach species were a very minor part of their collections.
In the mid-eighteenth century, Mauritius acquired Casuarina equisetifolia and Calophyllum inophyllum from Madagascar and Terminalia catappa from Bengal. In the early nineteenth century, Morinda citrifolia was imported from India and Desmodium umbellatum from an unspecified source. In the late nineteenth century, Pongamia pinnata was imported from the Seychelles. All of these have become naturalized as volunteers in natural coastal habitats on Mauritius, probably spreading around the island by floating seeds. Terminalia is also dispersed by the native fruit bats or flying foxes.
The story of Cocos nucifera is less clear. Coconut palms were reported on the Mauritius coast in 1598 by the first Dutch expedition. Whether they arrived on their own or were introduced by the Portuguese is unknown. In any case, coconuts are rarely found today along the Mauritius coast except where planted.
It is not immediately obvious why any of these species should have had to await human transport in order to reach Mauritius. All except Casuarina are known to be capable of long-range dispersal. Casuarina seeds survive immersion in salt water indefinitely but are not buoyant. They are believed to cross ocean barriers by rafting, particularly by floating pumice on which they have been seen germinating. Moreover, all these species were native on various other Indian Ocean islands, including the Seychelles, which lie in the same longitude as Mauritius (fig. 2).
The Seychelles were uninhabited when discovered by a British ship in 1609; wild coconut palms lined the beaches then. The archipelago remained uninhabited until colonized by French from Mauritius in the mid-eighteenth century, initially to exploit the wild coconuts. The first colonists also encountered coastal Casuarina, Calophyllum , and Terminalia groves. They were amazed at the size of the Terminalia trees, which they had known in Mauritius as saplings. Morinda and Pongamia were probably also naturally established
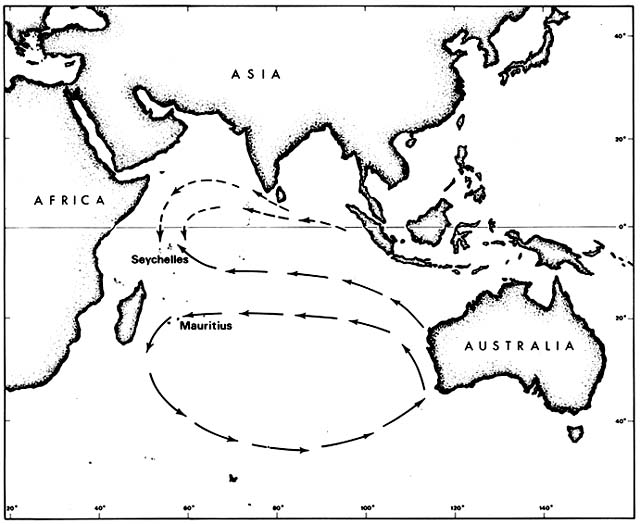
Figure 2. Prevailing Indian Ocean Currents Upstream from Seychelles and Mauritius. Although the currents
are not constant, drifting seeds of tropical beach plants are more likely to reach the Seychelles than Mauritius.
Some such species that arrived prehistorically in the Seychelles have since been artificially introduced to Mauritius.
in the Seychelles, although the first clear records are from the nineteenth century.
The rich native beach flora of the Seychelles, as compared to Mauritius, may result from the pattern of ocean currents. The Seychelles lie in the Equatorial Current downstream from Java, the rest of the East Indies, and many Pacific islands. Mauritius is far enough south to lie within the great counterclockwise circulation around the southern Indian Ocean and is thus downstream from the southern and western coasts of Australia. Such currents are variable, of course, but evidently not enough to erase different effects on dispersal.
Transfers Between Old and New Worlds
(Sauer 1982)
Some of the Indo-Pacific beach species introduced to Mauritius were also introduced through botanical gardens to the West Indies. Casuarina equisetifolia and Terminalia catappa from the East Indies were being grown in Jamaica by 1790, and Morinda citrifolia probably arrived soon after. All three were widely planted in the New World tropics during the nineteenth century, Casuarina for timber and fuel, Terminalia as a shade tree, and Morinda as a medicinal and dye plant. Herbarium specimens show that by mid-nineteenth century, all three had escaped from cultivation and were spreading on their own along seashores. Once in the region, Terminalia and Morinda were quite capable of island hopping by sea dispersal; both are recorded as having arrived in Grand Cayman, for example, as drift seeds. Casuarina and Morinda mainly colonize open seashore habitats, but Terminalia invades dense coastal thickets and overtops native trees.
Not all introduced Indo-Pacific seashore species have been so successful. Cocos nucifera, Barringtonia asiatica , and Scaevola taccada are commonly planted in the Caribbean region, often around beach houses, but are not spreading on their own.
Various Caribbean beach species have been widely planted in the Indo-Pacific region, including Mauritius, and have escaped to a few beaches, for example, spiderlily, Hymenocallis caribaea ; yellow bells, Tecoma stans ; whitewood, Tabebuia pallida ; frangipani, Plumeria spp.; and cocoplum, Chrysobalanus icaco . The cocoplum has become particularly successful on beaches of Viti Livu, Fiji. It had escaped to several beaches by 1962, and by 1984 it was extensively codominant with the native shrubs in beach ridge thickets on Viti Livu.
Anacardium occidentale , the cashew, has naturalized more extensively. Native to coastal dunes of northeastern Brazil, the cashew was introduced
by the Portuguese early in the colonial period to West and East Africa, India, and the East Indies; other people soon joined in planting the trees. The fruits are attractive to many animals, including monkeys, and the seeds are capable of at least local sea dispersal. Herbarium specimens show that by the mid-nineteenth century, the cashew was naturalized on many seashores of East and West Africa, tropical Asia, and the East Indies.
Comment
Like mangrove swamps, tropical sea beaches have a rich and very discrete flora. Few species transgress between these beaches and other kinds of habitats. There are minor exceptions, including the cases noted above when inland weeds temporarily invaded beaches after hurricanes and torrential rains. Conversely, beach species can invade adjacent inland sites when their normal competitors are removed. For example, in the Caribbean and Indian Ocean after World War II, abandoned military installations were commonly overrun by Canavalia maritima and Ipomoea pes-caprae vines.
For the outpost species, normal survival in a habitat within reach of storm waves involves perpetual micromigration, even on a stable coastline. Less extreme situations than the hurricanes discussed above include winter northerly storms and summer calms along the Mexican Gulf coast and monsoonal wind shifts on the East African coast. Beach ridge thicket species live in habitat largely controlled by a salt spray gradient and only marginally affected by storm waves. Where a coastline is progressively eroding or advancing, both outpost and thicket species may survive by local migration without any species extinctions within sites or recruitment of species from other sites.
Sea dispersal of tropical beach species has allowed them to colonize habitats that are spatially extremely discontinuous, not only on islands but on mainland coasts. Both outpost and thicket species are excluded from topographically sheltered shorelines by mangroves and inland species. Some beach species are excluded from bedrock or shingle coasts or from black sand or quartz sand beaches. Moreover, unlike mangroves, beach species need fresh water. They can survive a dry season by drawing on the fresh water lens floating in the sand above the denser salt water, but they are excluded from coasts where there is too little rain to maintain this lens.
Unlike many mangroves, the beach species produce seeds that remain dormant while afloat, with the embryos protected from seawater. As discussed in the Introduction, in two beach ridge thicket species, Tournefortia argentea and Scaevola taccada , breaking of dormancy requires a sequence of flotation in the sea, brief or long, followed by stranding and wetting by rain. Other beach species have seeds with extremely thick, hard integuments that remain impermeable during flotation for at least several months; they also
resist scarification by surf and sand. Presumably they germinate when the integument decays after long burial in moist sand.
As in the mangroves, adaptation for sea dispersal of beach species is probably not the result of selection for long-range dispersal per se, but for persistent buoyancy, since any seeds that sink at sea are eliminated as contributors to the gene pool of their species. The effectiveness of such selection is shown most dramatically by the outpost species, a dozen of which have naturally pantropical ranges. A few thicket species also have long-buoyant seeds and have achieved naturally pantropical ranges. Most thicket species have seeds that float for more moderate time spans or have fruits attractive to birds and bats, or both. It is within this latter group that recent human dispersal has permitted certain species to naturalize on beaches they had not been able to reach before.
Temperate Sea Beaches And Dunes
Mid-Atlantic Barrier Islands, United States
(Dolan et al. 1977, 1980; Leatherman 1979; Martin 1959)
A chain of barrier islands, with salt marshes and lagoons behind them, lies along much of the mid-Atlantic coast of the United States. Beaches are composed mainly of quartz sand, much more easily moved by the wind than the calcareous sand typical of tropical beaches. As a rule, dunes begin not far above the normal high water mark, leaving only a narrow, discontinuous storm beach at the top of the foreshore. This precarious habitat has a small but distinctive native herb flora. Some species are annuals, for example, sea rocket, Cakile edentula ; seaside spurge, Euphorbia polygonifolia ; and a rare beach pigweed, Amaranthus pumilus . Others are perennials, for example, beach pea, Lathyrus japonicus , and beach sandwort, Honkenya peploides . Only the Amaranthus is endemic; most are shared with beaches of the Great Lakes. Lathyrus and Honkenya are circumboreal.
The dunes form primarily under coarse rhizomatous grasses: in the north, beach grass, Ammophila breviligulata , shared with the Great Lakes and the Atlantic coast up to Newfoundland; in the south, sea oats, Uniola paniculata , shared with the Gulf of Mexico coasts as far down as Tabasco. Both are capable of virtually unlimited horizontal and vertical growth. Thriving under sand burial, they build high, steep dunes. These dune grasses grow in irregular clumps and patches, leaving many bare pockets and swales. Toward the dune front, these bare places have scattered individuals of the
same plants that colonize the storm beach. More protected swales and older dunes to the rear have a much richer flora. A few members of this flora are primarily coastal species: salt marsh cordgrass, Spartina patens ; beach plum, Prunus maritima ; bayberry, Myrica pennsylvanica ; beach heather, Hudsonia tomentosa ; and seaside goldenrod, Solidago sempervirens . These have wide coastal ranges outside the mid-Atlantic region and most also occur in scattered inland saline and sandy habitats. The bulk of the species present in the inner dune flora are primarily inland in distribution. Some are pioneers of open habitats, for example, little bluestem, Andropogon scoparius ; red cedar, Juniperus virginiana ; and poison ivy, Rhus radicans . In protected dune hollows, forests develop that have wind-shorn tops but share tree species with inland forests.
Prevailing winds are generally offshore, thus, traumatic events for the beach and dune vegetation are concentrated during storms. Occasional tropical hurricanes, with huge waves superimposed on storm surges, cause washovers and sometimes cut new inlets clear through barrier islands. Each year, 35 to 40 ordinary winter cyclones have onshore winds and waves strong enough to erode beaches. Occasionally one of these causes catastrophic erosion and washovers comparable to hurricane effects.
These barrier islands are believed to be inherently unstable and, in the long run, not under control of the vegetation. Large masses of sediment were moved landward during the great postglacial rise in sea level. About 6,000 years ago, as sea level began to stabilize at nearly its present level, waves, longshore currents, and winds working together formed barrier islands from the surplus sand. As long as the inshore zone contained sufficient sand, the islands built seaward, with the oldest, highest dunes at the rear. This progradation continued until about 2,000 years ago when the islands were much wider than now. Thereafter, net removal of sand offshore began; also blowouts and washovers pushed some sand over the islands into the lagoons. Thus, at present the islands are becoming narrower and migrating landward. As the islands become narrower, washovers are becoming more frequent, causing extensive burial by sand of the salt marsh vegetation along the lagoons. In the southern part of the region, marsh plants, including Spartina patens and Solidago sempervirens , commonly survive in the washover fans by upward regrowth. In the northern part, overwash fans are often colonized by Ammophila breviligulata , initiating a new, inner line of dunes. Remains of former inner dune forests are being exposed on some eroding foreshores.
Air photography is now being used to monitor shoreline changes over a 630-km reach of barrier islands. Transects at 100-m intervals, 6,300 in all, have been plotted spanning 15 years for the whole island chain and 30 years for much of it. Overall retreat averages 1.5 m/year, with more rapid retreat toward the northern ends of most islands and some accretion toward the southern ends. Retreating roughly 1 mile per century, changes are obvious
to residents. Inevitably, attempts to stabilize the islands are being made. A common approach is to develop continuous stands of dune-building grasses by planting, fertilizing, and watering. This commonly produces a higher, steeper foredune. With greater shelter from the foredune, vegetational succession at the rear proceeds toward an inland forest type. However, since the sand budget is not balanced, the steepened foredune is expected eventually to be undercut and washed away by storm waves, with complete washover of the islands following.
Mustang Island, Texas
In 1959, I laid out what were intended to be permanent 1-m-wide belt transects across the dunes of Mustang Island off Corpus Christi Bay. As usual on Texas barrier islands, there was a gently sloping intertidal beach, about 20 m wide. Just above normal high tide mark and at the base of the abruptly rising foredune there were a few widely scattered plants of Cakile geniculata and Amaranthus greggii , annual herbs endemic to Gulf of Mexico beaches on both sides of the tropics. The dunes were dominated by Uniola paniculata (discussed in the preceding section), joined here by the pantropical Ipomoea pes-caprae and the Caribbean Croton punctatus . On the backslope of the foredune were scattered grasses and dicot herbs, such as Spartina patens, Sporobolus virginicus, Oenothera drummondii , and Heterotheca latifolia . About 20 or 30 m behind the foredune, stabilized dunes had dense vegetation in which the outpost species were joined by Physalis viscosa, Rhynchosia americana, Ambrosia psilostachya , and a variety of other species. Nearly the whole assemblage was shared with the tropical Gulf coast. The transects were based on a government triangulation structure and were staked.
In 1961, the eye of Hurricane Carla crossed the coast close to the study site with winds of about 275 km/hr and waves reaching 7 m above ordinary high water mark. In 1962, the triangulation structure and stakes had disappeared and the transects could be only approximately relocated. Instead of being abut 20 m wide, the intertidal bare beach was now 80 to 90 m wide, extending clear through what had been the foredune zone and into the formerly stabilized dunes behind. Above a steep sand bank, freshly cut by the storm waves, the old dune vegetation appeared undamaged. At the top of the new beach and at the base of the cut bank, there were a few isolated seedlings of Sesuvium portulacastrum and Ipomoea pes-caprae and some resprouting rhizomes of Uniola and Spartina . Near the transected area, where washovers of barrier islands had occurred, seedlings of various beach outpost species were colonizing bared areas far back in the wrecked dunes, the seed presumably having ridden in on the storm waves.
Intercontinental Migration of Ammophila
(Cooper 1958; Dicken et al. 1961; Franklin and Dyrness 1973; McLaughlin and Brown 1942; Rosengren 1981; Sweet 1981)
Marram grass, Ammophila arenaria , native to western Europe, is even more vigorous as a dune builder than the eastern North American A. breviligulata (discussed above). Marram grass has been widely planted for dune stabilization. Where it has been introduced, it commonly has naturalized and spread.
On the Atlantic coast of the United States, marram grass is now naturalized locally, but has not generally displaced its native congener. On the Pacific coast, it has widely displaced a native dune grass, the circumboreal Elymus mollis , which is a relatively weak dune builder.
The amount of sand carried down western North American rivers was drastically increased during the nineteenth century by hydraulic mining, grazing, logging, and farming. Beaches near river mouths built out faster than vegetation could follow and surplus sand moved inland in great sheets and moving dunes. Starting in 1869, the active dune field that was to become San Francisco's Golden Gate Park was stabilized by planting marram grass. Soon after, the Coast Guard planted it to stabilize dunes along Humboldt Bay in northern California. Between 1910 and 1934, the Forest Service, other government agencies, and property owners planted marram grass on many dunes along the Oregon coast. Beginning in 1935, a huge dune field on the Clatsop Plains, south of the Columbia River mouth, was planted with marram grass. The eastern North American Ammophila breviligulata has also been planted on Pacific coast dune fields, but to a much lesser extent.
At the present time, Ammophila is thoroughly naturalized on innumerable stretches of the Pacific coast and has become a powerful geomorphic agent by building fairly continuous wall-like foredunes, which were not previously characteristic of this region. Sheltered behind these walls, formerly active sands have become vegetated with a mixture of native and exotic species. Prominent among these is Scotch broom, Cytisus scoparius , native to sands and sea cliffs of western Europe, which was also deliberately introduced in the late nineteenth century for sand stabilization and has now become thoroughly naturalized.
Marram grass has had a similar history in temperate Australia. Two native Australian beach grasses, Spinifex longifolius and S. hirsutus , are moderately effective sand binders, but they do not form steep, stabilized dunes as Ammophila does. Ammophila is currently replacing Spinifex hirsutus at latitudes above 33°S on many Australian beaches. Substituting stabilized linear foredunes for active transverse dune fields allows advance of woody inland
vegetation, especially Acacia spp. at the expense of various beach pioneer species.
Naturalization of Chrysanthemoides , Australia
(Gray 1976)
Chrysanthemoides monilifera is a shrubby composite native to the temperate southeastern coast of Africa, where it grows behind the storm beach on dunes and sea cliffs. The seeds are not buoyant and are probably bird dispersed. The species spreads vegetatively with vigor.
The species has been cultivated in the Sydney Botanical Garden and elsewhere in Australia since the late nineteenth century. Early in the present century, it escaped to beaches in New South Wales and on Lord Howe Island offshore. After 1950, it was much planted by the Soil Conservation Service to stabilize dunes and revegetate sand mining areas. This planting was stopped when it was found that Chrysanthemoides was invading and in places eliminating the complex native dune flora. In 1981, I saw several dense pure stands of this exotic on New South Wales beaches between 28° and 36°S latitudes, but none on any Queensland or Victoria beaches.
A reciprocal invasion of native South African coastal vegetation by introduced Australian Acacia cyclops and A. saligna is out of control (Taylor 1978).
Naturalization of Mesembryanthemum , California
(Blake 1969; Ferren et al. 1981; Moran 1950; Vivrette and Muller 1977)
The genus Mesembryanthemum s.l. (sometimes split into more than 100 genera) has thousands of species native to South Africa. Many of these are widely cultivated by fanciers of succulent plants. For example, M. edule (= Carpobrotus edulis ), native to the Cape Province and Natal, has been cultivated in Europe since the seventeenth century and in Australia and California since the nineteenth century. Although now widely naturalized there on sea beaches, it is generally recognized as an exotic that has escaped from cultivation.
Migration of a species very closely related to Mesembryanthemum edule , namely M. aequilaterum (= Carpobrotus aequilaterus ) is problematic. It is not known from South Africa and is generally considered to be native to either Australia, Chile, or California, or all three. The species was described in 1798 by Haworth from plants cultivated in England that were reported to be
"native to the country around Botany Bay." Either by oceanic drift or by transport on a ship, the species reached Chile before 1810, when it was collected at Valparaiso and named M. chilense . The first California record is from Bodega Bay in 1841. There are various plant species that apparently have naturally disjunct ranges between Chile and California, presumably due to dispersal by migratory birds. However, in this particular case, nineteenth-century human introduction appears likely. Mesembryanthemum aequilaterum is still actively spreading on the California coast. For example, at Point Dume in Los Angeles County, local residents have observed its expansion; in what was planned as a natural coastal reserve, it is spreading as a thick blanket that is overwhelming the native dune vegetation. However this species reached Chile and California, it seems that it originated in Australia along with several very closely related species that are Australian endemics. One of these, M. glaucescens (= Carpobrotus glaucescens ), appears in a water color painted at Sydney by Governor Arthur Philip about a year after the arrival of the first British colonists. The progenitor of these endemic Australian species must have come from South Africa long before, perhaps by oceanic drift.
Several other Mesembryanthemum species that are known to have been imported from South Africa have escaped from cultivation and are naturalized along the coasts of California, Baja California, and the offshore islands, for example, M. crystallinum and M. nodiflorum , which have been spreading since 1890, and M. croceum , which has been spreading since 1940.
Naturalization of Cakile , Australia and Western North America
(Barbour and Rodman 1970; Rodman 1974)
Various species of Cakile , the sea rockets, grow as extreme outpost pioneers on storm beaches. As noted in the Introduction, they have a peculiar mechanism for dividing their seeds between in situ reproduction and sea dispersal. Part of the seed is buried with the dead annual parent and part floats away, remaining viable for at least 10 weeks in seawater, but germinates only after being stranded and rained upon.
Cakile edentula is native to the Atlantic coast of North American from Labrador to Florida and the shores of the Great Lakes. In Australia, it appeared near Melbourne before 1863 and near Sydney by 1870, supposedly brought in sand ballast by an American sealing ship. It soon spread on its own to all the southeastern Australian states, including Tasmania, and to Lord Howe Island (fig. 3). By 1922 it was moving north along the temperate Queensland coast and by 1960 reached Heron Island on the Tropic of Capricorn. In 1981, I did not find it on any tropical Queensland beaches. Moving
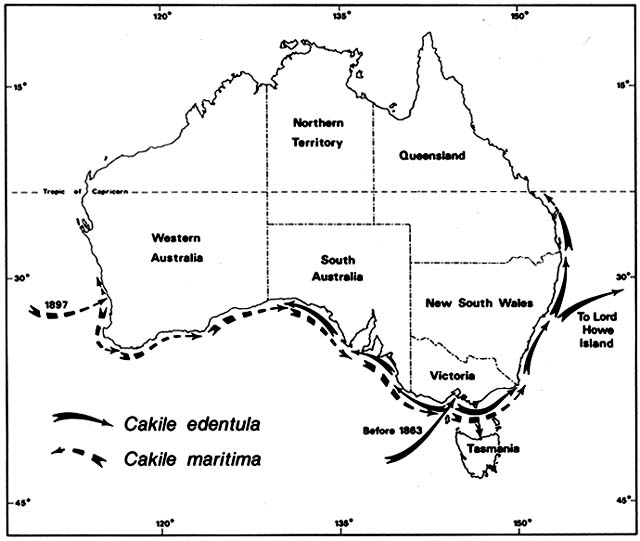
Figure 3. Naturalization of Cakile on Australian Beaches. Temperate beach species have repeatedly found open
niches distant from their native ranges following introduction by human agency. Two species of sea rockets that
arrived in Australia in the late nineteenth century spread by sea dispersal from their entry points along Australian
beaches, often advancing at a rate faster than 50 km/year until reaching their climatic limits. These grow closer to the
sea than native beach plants.
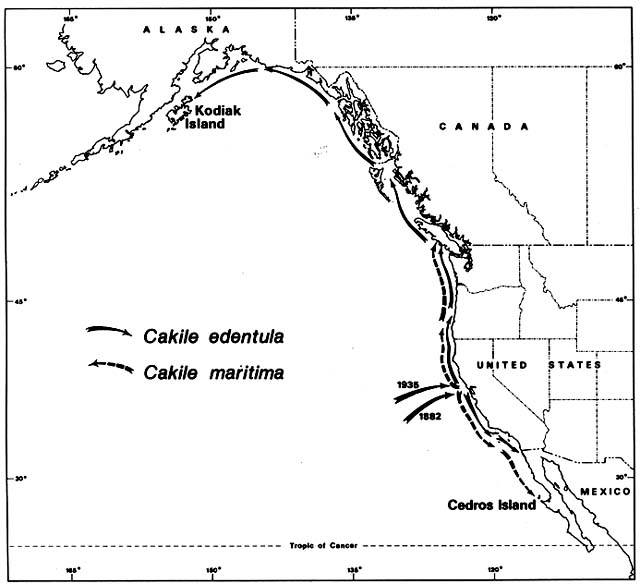
Figure 4. Naturalization of Cakile on Western North American Beaches. The sea rockets that invaded Australian
beaches were also accidentally introduced to the California coast. Cakile edentula from the Atlantic coast of North
America arrived first in San Francisco Bay. It then spread rapidly northward and southward. Before its migrational
history was investigated, some botanists mistook it for a native. Cakile maritima arrived much more recently from
Europe and has always been recognized as introduced.
westward from Melbourne along the south coast, C. edentula met and mixed with another invading sea rocket, C. maritima , which was moving in the opposite direction.
Cakile maritima is native to the Mediterranean and Black seas and to the Atlantic coasts of Europe as far north as southern Scandinavia and Scotland. It arrived at the port of Fremantle, Western Australia, by 1897. By 1963, it was ubiquitous on Western Australia beaches from 31°S southward; I found it on every one of 18 beaches studied in this region. Moving eastward along the south coast, C. maritima had entered South Australia by 1918 and Victoria by 1922. For a while, C. maritima and C. edentula shared the coasts of South Australia and Victoria, but C. martima now has sole possession of these Mediterranean type, dry summer coasts. Cakile edentula remains dominant along the temperate east coasts of New South Wales and Queensland, where summers are wetter.
On the Pacific coast of North America, the same sea rockets have immigrated in a pattern remarkably parallel to that in Australia. Again, Cakile edentula arrived first (fig. 4); the earliest record was on the Berkeley shore of San Francisco Bay in 1882. By 1891 it was common around the Bay, and outside the Golden Gate it had moved 30 km south to Half Moon Bay. By 1906 it had arrived at San Diego, where its southward expansion ended. By 1952 it was on the Channel Islands off the coast of southern California. Meanwhile, moving northward from San Francisco, it arrived in Oregon by 1901, Washington by 1907, British Columbia by 1909, and Kodiak Island off Alaska by 1931. The northward expansion had averaged 65 km/year over a 50-year period.
Local botanists, aware of the rapid spread of Cakile edentula , recognized it as an immigrant. However, the then dominant Harvard taxonomist, M. L. Fernald, temporarily confused the story by suggesting the Pacific coast populations belonged to a native endemic variety, which he named C. edentula var. californica . He postulated a formerly continuous boreal range, which became disjunct during the Pleistocene, a concept closely related to his nunatak hypothesis for other North American disjuncts. This variety is no longer considered distinguishable from the species proper.
The European Cakile maritima arrived in California so recently that no botanists have mistaken it for a native. It was first recorded in 1935 on a popular beach 25 km north of San Francisco. It spread almost as fast as C. edentula , averaging about 50 km/year in both directions. Northward it reached British Columbia by 1951, and southward it reached Cedros Island off the coast of Mexico by 1963. As in Australia, C. maritima almost completely replaced C. edentula in the region of Mediterranean type, dry summer climate. From Oregon northward, in areas with summer rain, C. edentula has remained predominant.
In both Australia and western North America, if only the present, static distributions of the two sea rockets were known, their nearly discrete ranges would appear to be directly controlled by climate. However, the actual history shows that Cakile edentula was quite successful outside its present climatic limits before the arrival of C. maritima . The outpost zone on a storm beach would at first glance seem one of the last places in the world where competitive displacement of one species by another would take place. Only a small part of the area is occupied by plants and for only part of the year. Nevertheless, if we recognize that the beach is not homogeneous, that there may be very limited safe sites for sea rocket establishment, and that the expanse of bare sand may be an uninhabitable wasteland for both species, we can visualize fierce competition in spite of low population density.
Comment
Temperate beach plants, like tropical ones, are relatively indifferent to rainfall gradients provided there is enough rain to maintain a freshwater supply under the sand. The greater seasonality of temperate coasts probably affects beach plants mainly via changing wave energy. Lacking fringing coral reefs, temperate beaches commonly have an annual cycle of strong erosion and accretion. Also, temperate zone beaches are generally composed mainly of quartz sand, which is more restricted in distribution in the tropics. Quartz sand is transported by longshore currents and is easily moved by the wind when dry, unlike calcareous sand, which is of local origin and normally immobile under the wind. Thus, many temperate barrier beach and dune systems are relatively extensive and dynamic. In spite of this, their floras are relatively poor in primarily coastal species except in South Africa and Australia, regions that are also phenomenally rich in primarily inland floras on ancient sand plains.
The primarily coastal, drift-dispersed outpost species have less broad and disjunct ranges in temperate than in tropical regions. A few that now have disjunct circumboreal ranges may be relicts from ancient distributions along former seaways. Similarly, the eastern North American beach species that now have disjunct ranges on the Great Lakes and Atlantic coast may have colonized the interior in early postglacial time when a marine embayment provided a continuous route, subsequently broken by isostatic uplift (Rodman 1974). Clearly, this species group is dependent not on salt water or salt spray but on waves, wind, and moving sand to create its habitat.
Like tropical beach pioneers, temperate outpost species need drift dispersal to occupy a dynamic and discontinuous habitat, but the temperate species have seeds that are buoyant for a shorter time. Perhaps this is because seeds
floating longer distances in temperate latitudes would have less chance of survival because the prevailing currents run north and south along the coasts rather than east and west through island stepping stones. The examples noted above of naturalization and spread following artificial introduction to distant regions demonstrate that the species had been unable to reach parts of their potential areas by natural long-range dispersal. They also demonstrate these species' capacity for rapid stepwise migration along a coastline and to offshore islands. Many similar cases, not discussed above, are known, for example, naturalization of Artemisia stelleriana and Carex kobomugi , native to northeastern Asia, on eastern North American beaches, and naturalization of Rumex frutescens , native to temperate South America, on British and Danish dunes. Some of these naturalized exotics have found an unoccupied niche closer to the sea than the native beach vegetation; others have invaded and displaced native dune species.
Salt Marshes
Limited Successions in Coastal Marshes, Britain
(Chapman 1959, 1976; Ranwell 1972; Tansley 1947)
Salt marshes have long been a favorite object of ecological studies in Britain; their history and dynamics do not support a model of succession leading to climax vegetation. Tansley (1947) concluded long ago that there is no good evidence that salt marshes can develop into nonmaritime vegetation by the mere accumulation of silt or humus.
A time-lapse study of a complex of Norfolk marshes over 25 years showed that in local areas where accretion of sediment happened to outrun regional subsidence, there was a predictable sequence of plant species. Commonly the first colonist of accreting mudflats, just above low tide mark, was a creeping monocot, Zostera noltii , followed in sequence by Salicornia europea, Aster tripolium, and Spartina townsendii s.l. The last is a rank, rhizomatous grass that is an exceptionally effective mud binder, at least temporarily; it is of hybrid origin and has only existed since about 1870.
There are many ways in which such autogenic salt marsh succession can be halted or reversed. Sedimentation may be offset by compaction so that there is little rise in marsh level. Accretion may shift rapidly to erosion because of a change in sediment supply or with strong onshore winds. Established marsh vegetation may be killed by deposition of flotsam and by
formation of stagnant pools and hypersaline flats. Also, marshes undergo cyclical deposition and dissection along tidal creeks. Steep banks inevitably develop along the creeks by a combination of deposition of natural levees under overbank flow on flood tides and scouring of channels on ebb tides. Eventually undercutting and collapse of banks starts the cycle over.
Limited Successions in Coastal Marshes, California
(Pestrong 1965; Warme 1971)
A similar cycle of bank buildup and collapse occurs along tidal creeks on San Francisco Bay mudflats. There is a correlated cycle of two dominant perennials, a pickleweed, Salicornia virginica , which grows on the natural levees, and a tall cordgrass, Spartina foliosa , which grows in the tidal creek channels, where it may endure 20 hr of submergence daily. As sediment is deposited on the natural levees at flood tides and scoured from the bank bases at ebb tides, banks become oversteepened and collapse. Salicornia does not survive submergence on the slump blocks but is replaced by Spartina , which traps sediment until bank buildup allows recolonization by Salicornia .
Salt marsh dynamics are more complex on Mugu Lagoon in southern California. Topographic maps spanning more than 100 years show that the eastern end of the lagoon, about 1.5 km long and as much as 0.5 km wide, has been mostly converted from open water to intertidal marsh. Recent measurements of staked areas show marsh advance at rates as high as 1 m/year. The marsh plants show fairly regular zonation, part of which can be fitted to an autogenic succession model. The subtidal lagoon has beds of eelgrass, Zostera marina , and ditchgrass, Ruppia maritima , which help trap sediment and shells and upgrade the substrate to the low tide mark. There is a barren zone above low tide mark where aggradation must be independent of plants. Spartina foliosa , which occurs at this level in other California marshes, is present here in only small, isolated patches. Above this bare zone, the lower fringe of marsh vegetation is dominated by annual Salicornia europea and perennial Salicornia virginica . Above 1.5 m above low tide mark, these chenopods are joined by 10 or so other species, mostly perennials, including other chenopods. These continue to the upper reach of highest tides, where they are joined by Arthrocnemum subterminale as a dominant. The vegetation of the middle and upper marsh is generally very thick and evidently capable of aiding sedimentation. However, rather than proceeding toward climax upland vegetation, older parts of the marsh become hypersaline areas, with salinities over twice that of seawater during the rainless summers, where expanding bare salt pans develop.
The Mugu marsh is strongly affected by various geomorphic processes that are not under control of autogenic vegetational succession. The marsh lies at the mouth of Calleguas Creek, normally a minor intermittent stream. During exceptionally heavy rains, the creek pours huge quantities of sand into the lagoon. The lagoon and marsh lie behind a barrier beach, the last of a series that formed sequentially offshore on an uplifted fault block since postglacial sea level stabilized. Sand being brought down the coast by longshore currents enters the lagoon through the tidal inlet. Also, during winter storms, ocean sand is poured over the barrier beach by washovers. A deep submarine canyon, with its headwall in unconsolidated sediments, has cut back almost to the mouth of the tidal inlet. It is on the verge of capturing the longshore sand supply and reversing the whole process of marsh development.
Primary Succession on a Mediterranean Lagoon, France
(Braun-Blanquet et al. 1958)
Soon after hearing about Clementsian succession theory, Braun-Blanquet initiated a time-lapse study to test the postulated relationship between salt marsh zonation and primary succession. The site chosen was on an aggrading tidal lagoon shore near Montpellier; initially it included a cove 30 m wide behind a narrow sand spit. The site was mapped nine times over a 43-year span; the study ended when the site was artificially disturbed. Seaweeds and marine angiosperms, for example, Ruppia maritima and Zostera marina , were present below low tide mark but were not mapped.
When the study began in 1915, the sand spit was mainly bare except for a narrow fringe near the high tide mark of annual chenopods, such as Salicornia europea, Suaeda maritima, and Salsola soda, and a few common reeds, such as Phragmites australis , that had taken root from stranded rhizomes. Along the shore of the lagoon was a belt a few meters wide dominated by perennial chenopods, such as Arthrocnemum spp. and Halimione portulacoides ; between them and higher ground was another narrow belt dominated by a rush, Juncus maritimus . Within its original area of about 100 m2 , the site also had 19 other less abundant species, all halophytes, such as sea lavender, Limonium vulgare , and sea aster, Aster tripolium .
By the end of the study in 1958, wave-deposited sand and shells had caused the shoreline at low tide mark to advance about 50 m, adding over 1,000 m2 to the study area. Fresh shell ridges had cut off two depressions within the former cove; the shallower one was covered with perennial Arthrocnemum spp.; the deeper one retained a pool of open water inside an Arthrocnemum ring. The thin fringe of annual chenopods and reeds had moved forward with the advancing shoreline. Behind it, better-drained sand
and shell areas were dominated by a steadily expanding Halimione colony. At the rear of the study area, the initial Juncus zone had remained relatively stationary, but had moved slightly seaward into the Arthrocnemum -dominated area and was in turn being invaded at the rear by Spartina versicolor , a tall grass growing in both saltwater and freshwater marshes. At the same time, the better drained Halimione -dominated areas were beginning to be invaded by tall Juncus acutus and Limonium vulgare .
Interrupting the steady seaward advances were setbacks, when strong mistral winds and waves opened up patches of established vegetation for recolonization by the pioneer fringe species. Also during the rainy season, ephemeral populations of weedy annuals appeared on the tops of shell ridges, including Polygonum aviculare, Stellaria media, Galium aparine, Sonchus oleraceus, and Senecio vulgaris .
Measurements of soil variables and plant osmotic pressures suggested that species distributions were largely controlled by soil moisture, salinity, and aeration. Halimione occupied sites with the greatest contrasts between rainy and dry season soil conditions and had the greatest range of osmotic pressure, from less than 25 to more than 60 atm.
As for testing the model of primary succession, the results are equivocal. The results generally agree with the model in showing a tendency for the vegetation zones to advance as sediment was deposited on the shore. The results do not prove the crucial assumption of a Clementsian autogenic succession model, namely, that changes in the ecosystem are directional and under control of the vegetation. In this case, the changes may have been allogenic, that is, due to surplus shell supply, leaching of sediments by rain, or other processes not under vegetational control. The study site was chosen on an aggrading shore. If vegetation changes passively follow shoreline changes, the zones may be static or retreating on other shorelines.
Natural Colonization of Artificial Inland Salt Marshes,
Europe and North America
(Ogle 1981; Simon 1958; Zumph and Rebmann 1932)
As a waste product of salt mining operations, saline ponds were created 40 km south of Berlin in 1907. Within 3 years the ponds were colonized by halophytes, including Triglochin maritima, Glaux maritima, and Plantago maritima . The botanists who studied the colonization believed the seed had been brought from seacoast marshes by waterfowl; it is also possible the seed had been transported from other saline inland places, although none were known closer than 30 km.
Commercial exploitation of a salt dome at Müllhausen, Alsace, began in
1904. Salt ponds created by slurry dumping were visited by migratory waterfowl. By 1954, the ponds had been colonized by halophytes, including Chenopodium botryodes ; the nearest seed source known for some of the species was 500 km away.
In western Virginia, 400 km from tidewater, pumping of brine wells since the late nineteenth century has created a habitat for various salt marsh species, for example, Salicornia europea and Eleocharis parvula . A new species, Scirpus robustus , was found in 1977; its population expanded rapidly in the next few years. All the species are believed to be dispersed by birds.
Naturalized Exotics, California
(Robbins 1940)
In the late nineteenth century, E. W. Hilgard, head of the California College of Agriculture, imported seed of various species as possible forage plants for alkali lands. From Australia, Baron Ferdinand von Mueller sent seed of several species of Atriplex , including A. semibaccata , which has grown successfully at the Tulare experiment station and from there has been widely distributed in California. The first escape to a tidal marsh was reported by botanist Alice Eastwood in 1901. She found it flourishing in a marsh north of San Francisco. By 1935, it was widespread along the mainland coast and on offshore islands. Today it is extremely common, even dominant, in the upper parts of coastal marshes and other saline habitats.
Several other salt marsh species arrived in California by unrecorded means, an obvious possibility being in sand used for ship ballast. Brass buttons, Cotula coronopifolia , native to South Africa, was found in San Francisco Bay in the Early 1850s and has since become common along the whole mainland coast and on offshore islands. Two grasses native to western Europe and the Mediterranean, Monerma cylindrica and Parapholis incurva , were first recorded near San Diego about 1900 and have since spread far northward along the California coast; Parapholis has also reached offshore islands. A European chenopod, Bassia hyssopifolia , may have arrived in California indirectly via Argentina as a contaminant in alfalfa seed. It was naturalizing in inland alkali and saline habitats by 1920; at present it is invading coastal marshes near Santa Barbara.
Comment
A few exceptional plant species, including Phragmites australis and Cotula coronopifolia are shared between saline, brackish, and freshwater marshes, but
on the whole, salt marshes have a very discrete flora. Most salt marsh species are confined either to coastal marshes subject to tidal flooding or to interior marshes that may be only seasonally flooded, but a few are capable of growing in both. For species capable of coping with the extreme osmotic pressures of these habitats, the reward is escape from competition from ordinary nonhalophytes.
In many salt marshes, plant species can survive indefinitely with only micromigrations and no capacity for long-range dispersal. There are cases of salt marsh species that have evolved as localized endemics, for example, several Atriplex species in the San Joaquin Valley of California. However, many salt marsh species have very wide and disjunct distributions. Some species are known to be capable of long-range dispersal by ocean currents, such as Salicornia spp., which have seeds capable of floating for months (Ranwell 1972). Such dispersal may explain some grand disjunctions, such as the transatlantic ranges of Salicornia europea and S. virginica . It cannot explain the presence of these two species on both Atlantic and Pacific coasts of North America. For many such disjunctions, particularly involving inland populations, bird dispersal seems to be the best explanation. Disjunct salt spring floras in the interior of Europe have sometimes been attributed to fragmentation of populations that were continuous during Pleistocene arid periods, but Aellen (1958) argued that known cases of recent bird dispersal obviate the need for such explanation.
The cases noted above of naturalization following long-range human dispersal involved disjunctions that birds were unlikely to span.
Freshwater Aquatic Habitats
Vernal Pools, California
(Griggs 1981; Jain 1976; Keeley and Keeley 1979; Thorne and Lathrop 1969)
Seasonal lakes and pools in California grasslands are tiny relicts of native herbaceous vegetation in a sea of European annuals. They harbor over 200 species of flowering plants in about 50 genera, many of which are found in no other habitat. Most are in the Central Valley on Pleistocene alluvium underlain by an impervious hardpan or claypan. Some are on ancient lateritic soils on mesas formed by basalt flows.
Under the present Mediterranean climate, habitat conditions vary drastically with the seasons and from year to year. Most of the species are annuals.
These may bank large quantities of seed in the soil in wet years; the seed may remain dormant for years or decades. For example, in one pool a rare grass, Orcuttia pilosa , grew abundantly one year, disappeared completely for 4 years, and reappeared in abundance the fifth year, when the soil still contained 50 dormant seeds for every one that germinated. Orcuttia seed germinates only when infested with certain fungi, which are abundant only when the pond is flooded. Orcuttia seedlings start life as submerged aquatics, then develop floating leaves and become tall emergents when the water level drops; when the pool dries, flowering begins within days.
Annual censuses of the flora of a vernal marsh on the Mesa de Colorado in southern California showed species appearing that had not been seen in previous years, for example, Potamogeton pusillus and Elatine californica in 1968 and Orcuttia californica and Myosurus minimus in 1969. Whether these were new immigrants or from dormant seed is not known.
Some vernal pool species are endemic to California or at least to the far western United States, including several congeners in Orcuttia, Limnanthes, Elatine , and Downingia . Some are disjunct between temperate North and South America, for example, Deschampsia danthonioides, Elatine chilensis , and Lilaea scilloides . Some are shared between North America and Eurasia, for example, Crassula aquatica, Eleocharis palustris, Callitriche palustris, Potamogeton pusillus , and Myosurus minimus . A few range from the Old to the New World and from the northern to the southern hemisphere, such as Juncus bufonis and Lemna minor .
The likelihood of long-range bird dispersal of these species is discussed at the end of this chapter. Local bird dispersal of recently introduced exotics appears likely in a few cases. Naturalization of the South African Cotula coronopifolia in California salt marshes was noted in the preceding chapter. This species has also spread inland around vernal pools, including those on the Mesa de Colorado. Another annual, Lythrum tribracteatum , native to southern Europe and the Near East appeared in California in 1930 in a vernal pool near Sacramento. By 1948 it had spread 200 miles southeast in the Central Valley to a lake that dries each year; other widely scattered populations have since appeared in seasonally flooded central California sites. In 1978, Aponogeton distachyus , the Cape pondweed from South Africa, was discovered in vernal pools on Kearny Mesa in San Diego. The species is widely planted in ponds as an ornamental; it has also escaped to natural freshwater habitats in northwestern Europe, western South America, and southern Australia.
Many California vernal pools have quite recently been destroyed by conversion to rice paddies or drainage for other crops. Preserving a few relicts of this increasingly disjunct habitat type is currently an issue in California.
Intercontinental Migrations of Elodea and Hydrilla
(Mitchell 1977; Sculthorpe 1967; Yeo and McHenry 1977)
Two closely related waterweeds have invaded streams in various continents following long-range human introduction. Both are dioecious perennial herbs, rooting in sediment and with the submerged stems bearing whorled foliage like a bottlebrush. Flowers of the pistillate plants open at the water surface to receive floating pollen. Both species are easily propagated vegetatively and have become favorite freshwater aquarium plants.
Elodea canadensis , native to lakes and slow streams of North America, arrived in the British Isles in the 1840s, supposedly imported accidentally with American water lily rhizomes. Although only female plants were present initially, the species spread rapidly by human and perhaps bird dispersal. It was widely naturalized in the British Isles and on the continent by 1879, when a male clone arrived and seed production began. For some years, Elodea proliferated excessively, impeding water flow and boat traffic; control efforts were futile because of rapid regeneration from fragments of the plants. After a few years, populations generally declined spontaneously, possibly due to buildup of predators. Since 1900, Elodea has subsided to become merely another member of Europe's freshwater flora.
In Australia, Elodea escaped by 1960; its spread is unclear because for a while it was confused with the native Hydrilla . It has become a common weed of rivers and ditches in the New South Wales—Victoria border region.
Hydrilla verticillata is regarded as native in temperate and tropical eastern and northern Australia, the East Indies, southeast Asia, and various western Pacific and Indian Ocean islands. It can reach the surface and establish a dense canopy in water as deep as 15 m. Introduced to North America in the aquarium trade, it was first noticed growing wild in Florida in 1958; by 1967 it occupied tens of thousands of acres of waterways in Florida and neighboring states. Only male plants are present there, but the species is dispersed vegetatively, probably by waterfowl and trailered boats. Drag lines haul it out by the ton to keep waterways open, but it promptly grows back.
In California, Hydrilla was found to have become naturalized in the Sacramento Valley in 1976; it has since spread widely in the state. It has become a serious problem in canals and irrigation ditches in southern California. In 1985, the state Department of Fish and Game authorized the Hydrilla Control Research Project to stock all canals of the Imperial and Coachella valleys with 78,000 Amur River grass carp. A mutant triploid strain of the fish is being used, which is expected to live about 5 years and die without reproducing.
Hydrilla has escaped in many other warm regions in both natural and artificial lakes and streams. Gatun Lake, Panama, has a flourishing infestation.
A similar case of naturalization following long-range dispersal through aquaria involves Lemna minuscula , a tiny floating aquatic native to the New World; it has apparently been spread by bird dispersal through European lakes and streams (Landolt 1979).
Intercontinental Migrations of Eichhornia and Alternanthera
(Bock 1968, Hockley 1974; Maddox et al. 1971; Mitchell 1977; Russell 1942; Varshney and Rzoska 1976; Vietmeyer 1975)
Water hyacinth, Eichhornia crassipes , and alligator weed, Alternanthera philoxeroides , are native to the great river systems of eastern South America from the Orinoco to the Paraná. Both root in shallow water along stream banks and may extend out over deep, quiet water as floating mats. Both are capable of rapid vegetative reproduction. Water hyacinth has nitrogen-fixing bacteria within the leaf and grows well in clean water. Alligator weed grows best in nutrient-enriched water; it can tolerate brackish water better than water hyacinth. In their native habitats, neither species is especially abundant or conspicuous. When introduced abroad, both have commonly produced prodigious biomass, presumably because of release from natural predators.
At home or abroad, both species survive devastating floods. When the bulk of the floating mats are carried off, survivors in backwaters remain. Water hyacinth also stores dormant seed in the mud, which remains viable for at least 15 years. Outside its native area, alligator weed seems to reproduce only vegetatively; although it has bisexual flowers, a clone may be incapable of self-fertilization.
The long-range migrations of water hyacinth are easily explained by deliberate human transport; it produces beautiful flowers and shiny foliage. Alligator weed is an unattractive plant with no known record of deliberate planting. In spite of their differences, the two have had rather similar macro-migrations.
Water hyacinth was introduced from the Orinoco region to New Orleans, Louisiana, in 1884 and spread rapidly as a planted ornamental in pools and ponds all over the southeastern United States. Alligator weed arrived by unknown means in Florida by 1894 and in Louisiana by 1897. These two exotics have profoundly changed the geomorphic processes of the Mississippi Delta and other southeastern waterways. They are more indifferent to water depths than the native aquatics and produce much greater biomass. Bottom sediments under masses of these species are much higher in organic content
and nutrients and lower in oxygen; the whole economy of the water bodies is affected. Moreover, the result is not permanent filling of bayous and marshes with organic sediment that would initiate autogenic succession; the accumulation of weeds and sediment is eventually ripped out by floods.
Expensive efforts to control these species by mechanical or chemical attack have had only brief effect. Alligator weed is exceptionally tolerant of herbicides; herbicides kill water hyacinth plants but not the buried seed. Since 1960, several insect predators found on these weeds in South America have been released in the southeastern United States and are evidently having some effect.
Relatively minor colonies of both species have become established in California. Starting in 1904, water hyacinth spread gradually along the Sacramento and San Joaquin rivers and their tributaries. It has also established a few colonies in southern California, but there are no extensive suitable habitats there. Since 1965, alligator weed has colonized some artificial reservoirs and effluent ponds in southern California. Release of insect predators has been tried; ironically, the weed populations are so small and localized that the insect predators cannot maintain viable populations.
In Australia, water hyacinth has been widely planted since 1894. In New South Wales, it escaped to become a temporary nuisance along some rivers but has been nearly eliminated since 1935. In Queensland near the Tropic of Capricorn, construction of a barrage in 1970 on the Fitzroy River estuary greatly encouraged water hyacinth by stopping incursions of tidal seawater and flushing by freshwater floods. By 1973 the lower Fitzroy was covered with water hyacinth for over 50 km above the barrage. A costly spray and cutting program was followed in 1975 by release of a South American weevil obtained from a biological control program used in the southeastern United States. Alligator weed arrived in New South Wales by 1945, being first noticed on heaps of ship's ballast dumped near Newcastle. It has subsequently colonized about 10 localities, mainly in irrigation canals and rivers with sewage effluent.
In Java, water hyacinth was grown in the Buitenzorg Botanical Garden in 1894 and soon escaped to streams flowing through the garden. In Bengal, it was introduced from Australia in 1897. By shortly after 1900 it was being widely planted in Ceylon, Malaya, Siam, Indo-China, and China; its spread was partly in botanical gardens, largely in private hands. Its flowers were used as temple offerings, and it was planted in farm ponds to produce pig fodder, feed fish, and provide green manure. It has escaped to the Ganges, Mekong, and other rivers where it interferes with navigation and aggravates floods during cyclone passages. It chokes many of the canals in Bangkok. The alligator weed is rather uncommon in the East Indian—Southeast Asian region, although it arrived in Java by 1895 and has reached the mainland.
In Africa, the earliest available record of water hyacinth is from El Gizeh near Cairo, where it was growing in ditches and ponds in 1930. Evidently the lower Nile region offered little suitable habitat because the great invasion came by another route. In 1954, water hyacinth escaped from ornamental plantings in the Belgian Congo; it rapidly exploded into a major pest, interfering with navigation on the Congo River. In 1957, it was discovered in a remote part of the Sudan 300 km upriver from Khartoum on the White Nile. The watershed between the Congo and White Nile is narrow; perhaps cranes or other water birds carried the seeds across. In 1958, a survey found that the species was established along hundreds of kilometers of the White Nile; by 1962, it extended from Juba to the Gebel Aulia dam near Khartoum, a huge expanse bracketing the Sudd region. The Sudd is an enormous freshwater marsh, dominated by emergent aquatics, such as Cyperus papyrus and various species of Typha and Phragmites . Maintaining navigation channels through this morass had required heroic efforts; invasion by water hyacinth frustrated these efforts and, by raising the water level of the marsh and increasing evaporation, reduced the whole flow of the Nile. Work was begun on a 350-km-long canal to bypass the Sudd and increase the Nile flow, but has been halted by a long civil war.
Comment
Freshwater aquatic habitats harbor a rich and discrete flora. Whole genera and even families have evolved in these habitats. Monocots are especially well represented, but there are a variety of dicots and some gymnosperms. Growth forms range from the tiniest of herbs to great swamp trees. As noted in the preceding chapter, only a very few species are shared between freshwater, brackish, and salt marshes.
Geographic patterns of species are heterogeneous, but in general, microdistributions are sharply limited by water depth and other local variables while gross ranges are diffuse and disjunct. In the long run, survival of disjunct populations in dynamic habitats requires constant colonization. Local populations are subject to extinction by both catastrophic habitat change and by autogenic succession in low-energy habitats. In the classic theory of primary succession, Kerner von Marilaun (1863) postulated how a lake becomes a marsh and a marsh becomes a meadow.
Disjunctions in ranges of freshwater aquatics are so great that they have sometimes raised doubts about being caused by natural dispersal. Scirpus californicus , for example, has a highly disjunct range from California to Patagonia, Easter Island, and Hawaii. Heyerdahl (1971) suggested that this reed had been planted in the crater lake on Easter Island by prehistoric South American Indian voyagers who arrived on balsa rafts made from the reed. Howevever, growing in the same Easter Island lake is a species of no known
use, Palygonum acuminatum , which also has a widely disjunct range in the Americas. Heiser (1978a ) argued that bird dispersal of both species was more likely. Study of fossil deposits in Easter Island lake may determine when Scirpus arrived on the island.
Wind dispersal may be responsible for moderately wide disjunctions in some species, such as Senecio spp., and for extremely wide ones in others, such as the cosmopolitan cattails, Typha spp., and the common reed, Phragmites australis .
For most freshwater aquatics, migratory birds appear to be the most important agents of long-range dispersal. Ridley (1930) summarized a wealth of evidence that ducks and other water and shorebirds disperse aquatic plants. Some of the evidence is circumstantial, for example, colonization of isolated ponds and marshes that are visited by waterfowl and lack of colonization of those that are not visited. Much of the evidence, however, is quite direct, including many cases where shot waterfowl had viable seeds and vegetative parts in mud on their feet, adhering to their plumage, or carried internally. More recently, deVlaming and Proctor (1968) fed captive waterfowl seeds of 23 species of aquatic or semiaquatic plants. They found that viable seeds of most species were passed by killdeer or ducks or both after 20 hr or more, long enough for the birds to fly over 1500 km. Viable seeds of Potamogeton pectinatus were retained for 75 hr by both killdeer and mallards, and viable seeds of Eleocharis quadrangulata were retained 120 hr by killdeer.
Some wide disjunctions of aquatic plants fit neatly within major flyways of migratory waterfowl. The southern water hemp, Amaranthus australis , has its main populations in the Everglades and other marshes of the southeastern United States. Its seeds, ripening in the fall, are a major food of migratory birds taking off southward over the Gulf of Mexico and the Caribbean. Distribution of this species south of the United States is extremely disjunct, always in freshwater lakes and marshes: five localities in five different states in Mexico, seven localities in four West Indian islands, one lake in Venezuela, several rivers in the Guianas, and one locality in southern Brazil, which is 4,000 km from the closest Guiana locality but on the same flyway (Sauer 1972). Kloot (1984) noted that nearly all the plants shared between the pre-European flora of temperate South Australia and other continents, mainly Eurasia, are aquatic or marsh species that presumably were introduced from the northern hemisphere by migratory waterfowl.
In North America, slough grass, Beckmannia syzigachne , has evidently been dispersed northward by waterfowl. It has a disjunct but fairly coherent range in marshes of the north central and western United States and in the Canadian prairies, where its seed heads are fed upon heavily by waterfowl moving up the Central Flyway. Farther north its distribution is sporadic. In an intensive study of the arctic Mackenzie River Delta, Gill (1974) found Beckmannia only on lakes frequented by waterfowl and, on those lakes, only in places with especially heavy concentrations of waterfowl.
East—west disjunctions of aquatics between North America and Eurasia are less obviously related to flyways, but there are many records of birds breeding in the Nearctic but wintering in Eurasia and birds breeding in the Palearctic but wintering in the Americas. An example of the latter is the Eurasian widgeon, which breeds in the Palearctic from Iceland to Siberia but has been recorded as a casual visitor over much of North America (Edgell 1984).
The same species of Beckmannia discussed above is found in eastern Asia, the exchange presumably via Alaskan waterfowl breeding grounds. Heslop-Harrison (1953) suggested a similar relay of a number of aquatic plant species shared between eastern North America and Britain; waterfowl from both regions converge on the same summer breeding grounds in Greenland.
The cases of recent naturalization following human dispersal show that, however great the radius of natural aquatic plant dispersal, it was not global.
Riparian Habitats
Mackenzie Delta, Canada
(Gill 1973a,b )
In its great alluvial delta, the complex streams of the Mackenzie River constantly erode banks and destroy old vegetation on the outside of their shifting bends. At the same time, they create new ground for plant colonization on point bars deposited on the inside of their shifting bends. The details of erosion and deposition are complicated by freezing and thawing of the river and of the ground above permafrost and by flooding and ebbing of the river. However, accreting point bars have a fairly consistent sequence of colonization. The first pioneers are typically a nonseed plant, Equisetum , and Salix alaxensis , both capable of surviving regular flooding and rapid sediment deposit. After the site builds up above normal flood level, about 3 to 4 years later, these are joined by seedlings of Populus balsamifera , which for a while is the sole dominant tree. Under the canopy of this poplar, Alnus crispa and several Salix spp. form a shrub layer. The herb layer includes Hedysarum alpinum and several Arctostaphylos and Artemisia spp. The Alnus and Hedysarum have symbiotic nitrogen-fixing bacteria. Eventually, on the uppermost natural levee surface, which is only rarely flooded, Picea glauca invades and becomes the sole dominant as Populus dies out. The other pioneers remain in the understory of the spruce forest and are joined by many shade-tolerant herbs.
An almost identical succession is reported along accreting banks of Alaska floodplains (Van Cleve and Viereck 1981).
Meramec River, Missouri
(Dietz 1952; Sauer 1952)
The Missouri Botanical Garden Arboretum at Gray Summit is largely covered with an Ozark oak—hickory forest, but the Meramec River floodplain has a different kind of vegetation. Every spring the Meramec rises over its banks and changes the topography of its floodplain by erosion and deposition. Old maps and air photographs from eight different time levels over a 100-year span showed that the floodplain vegetation was generally less than 30 years old before being recycled. Pioneer colonization was dominated by Salix interior and other willows. Willow seed floating down the river on the spring flood is viable for only a few days and becomes established only if stranded on fresh alluvium while the flood is ebbing. In the area studied, willow seedlings had formed new rows in only 4 of the preceding 10 years. After 1 year, willows are too deeply rooted to be easily dislodged by the next flood. On accreting sandbars, older willow thickets are invaded by Populus deltoides, Ulmus fulva, U. americana , and Acer saccharinum , which soon become dominant. Other common riparian trees are Platanus occidentalis and Celtis occidentalis . All these trees have seeds dispersed by wind and water.
In addition to the riparian trees, other growth forms colonize sites opened by the river. Sunny sand and gravel bars have clones of switchgrass, Panicum virgatum , a coarse perennial with creeping rhizomes that are very resistant to erosion during floods. Other fresh alluvium unoccupied by trees has patches of rank annual herbs, such as giant ragweed, Ambrosia trifida , and rough pigweed, Amaranthus retroflexus ; both germinate from buried dormant seed when the substrate is disturbed. In a single growing season, one individual of either species may produce tens of thousands of seeds, some of which survive burial in the soil for decades until the soil is again disturbed.
A different colonizing pattern is followed by pokeweed, Phytolacca americana . Like many pioneers, it only germinates in disturbed soil. However, it grows well in both sun and shade and, once established, is not easily eliminated by competitors. The perennial root sends up new shoots every year for 30 to 40 years. The berries attract birds, which disperse the hard seeds widely; the seeds may remain dormant in the soil for decades until disturbed. Pokeweed is not restricted to stream banks, but they probably were its evolutionary cradle.
I mapped the distribution of individual pokeweed in a 500 by 1,500 m study site on the arboretum's Meramec floodplain and adjacent bluffs in 1947
and 1950; the same area was remapped by Alan Covich in 1962 (unpublished data). In 1947 the floodplain part of this site had 175 pokeweed plants, some in areas that had been recently logged, but many others in places with no trace of human activity. In the next 15 years, most of the original plants were carried away by flood erosion or crowded out by competition, so that only about a dozen of the original plants survived in 1962. During the same period, nearly 100 new seedlings became established, mainly in freshly disturbed sites. Such perpetual micromigration is probably the normal way of life for riparian species.
San Pedro River, Arizona
(Zimmerman 1969)
Arid southwestern North America has riparian vegetation remarkably similar to the humid east, considering their quite different regional vegetation. Some eastern species, including pokeweed, drop out west of the 100th meridian, but others, including Panicum virgatum , do not. As a rule, the eastern species are replaced by closely related congeners. In the Sonoran Desert of Arizona, the riparian annuals include Ambrosia aptera and Amaranthus palmeri ; riparian trees include Populus fremontii, Platanus wrightii , and Salix spp. Unlike the adjacent upland desert species, which have seedlings established mainly during summer rains, the Arizona desert riparian trees behave like their eastern congeners. Their seedlings are established on substrates wetted by spring snow melt, and they colonize water courses with sustained flow, not summer flash floods. The same is true of some desert riparian shrubs, such as the seep willow, Baccharis glutinosa .
A separate group of riparian species in the desert region is negatively associated with regularly flowing streams and concentrated along washes with ephemeral flows and no groundwater supply. An example is Chilopsis linearis , a tree with seeds that germinate when buried in warm, wet sand after a summer flash flood.
Tamarix and Other Riparian Invaders,
Southwestern North America
(Everitt 1980; Harris 1966; Robinson 1965; Turner 1974)
The tamarisks or salt cedars include various species of shrubs and trees native to riverbanks and wet saline places of the Mediterranean region and
arid parts of Eurasia. Beginning early in the nineteenth century, various Tamarix species were imported by United States nurseries and widely planted for their graceful feathery foliage and pink flowers. The different species are seldom separately identified by American laymen or botanists, so the tamarisk invasion has to be dealt with on the genus level. In this chapter, only the invasion of natural riparian habitats will be considered; the effect of dam building and altered river regimes will be discussed in Chapter 4.
Herbarium specimens record sporadic naturalization along rivers in the Southwest after 1875, for example, on a sandbar in the Gila River, Arizona, in 1898, but occurrences were local until after 1920. As late as 1926 the U.S. Department of Agriculture was experimenting with planting tamarisk along the banks of the Rio Grande for erosion control. Before 1930, the main tamarisk invasion was under way, and during the 1930s came general awareness from Texas to California of its arrival as an aggressive invader.
The greatest tamarisk infestations in natural habitats in the Southwest are along major rivers that flood from snowmelt in late spring and early summer. Like native riparian trees and shrubs, tamarisk produces seeds that are only briefly viable. Flowering and fruiting, however, extend from late spring through summer, so viable tamarisk seed is blowing in the wind and floating in streams in normal flood season. Tamarisk seeds can germinate while floating. Their best chance of establishment is when they are stranded on fine, fresh alluvium that remains moist enough for rooting.
The native Baccharis glutinosa , tamarisk's most direct competitor, has an advantage in producing seed all year, and if spring melt is early, its seedlings get a headstart. On the Gila and San Pedro rivers, Arizona, however, tamarisk eventually invaded established Baccharis thickets and took over. Tamarisk thickets also prevent establishment of new seedlings of Populus .
Along streams in the southwestern United States and northwestern Mexico that are dominated by summer flash floods, tamarisk has remained a relatively minor component of riparian vegetation. This may not be due directly to the seasonal difference but rather to the coarser sediments and rapid drying. However, scattered tamarisks have colonized sandy washes.
Tamarisk has recently jumped about 40 km of ocean to colonize Santa Cruz Island off the southern California coast. Starting in 1977, isolated tamarisk bushes have been found every few years at various places on the island. Each time the tamarisk was destroyed without being allowed to produce seed. The nearest known seed source is at stream mouths on the mainland coast near Santa Barbara. It seems likely the seeds are blown across the channel by the occasional strong northerly winds.
Several other exotic species deliberately introduced into the southwestern United States have escaped and spread widely on streambanks; Russian olive, Eleagnus angustifolia , and Siberian elm, Ulmus pumila , are examples.
The coastal mountains in southern California have a fairly rich native riparian flora, although few of the streams are perennial. Gallery woodlands are dominated by deciduous species of Alnus, Populus, Salix, Platanus, Acer , and Juglans and by evergreen Quercus and Umbellularia . Understory species include Baccharis glutinosa and Artemisia douglasiana . Yet even where free from human disturbance, this apparently well-stocked community is commonly invaded by exotic shrubs, especially the African castor bean, Ricinus communis , and the South American tree tobacco, Nicotiana glauca . The castor bean is believed to have been introduced during the Spanish colonial period as a source of oil for lamps and medicine. The tree tobacco is native to riverbanks, gullies, and rocky cliffs in the Chaco and Mato Grosso, but may have come to California via Mexico, where it is widely naturalized. The earliest available California record is from 1879 at Los Angeles. German ivy, Senecio mikanioides , native to South Africa, is currently festooning some southern California riparian woodlands with masses of vines.
Riparian Invaders, Australia
(Buckley 1981; Meadly 1954)
In arid central Australia, few alien plants have penetrated most habitats, even along tracks of oil exploration. Water courses are the main routes of invasion. Just as in California, Ricinus communis and Nicotiana glauca are naturalized on stream banks; other aliens include a European sowthistle, Sonchus oleraceus , and a South American cocklebur, Xanthium spinosum . All of these had arrived in coastal Australia during the nineteenth century.
A prickly poppy, Argemone ochroleuca , native to arroyos of Mexico, had arrived in New South Wales by 1845; it has since spread widely, mainly as a weed but also along streams. In 1981, I saw thriving colonies of it along a natural stream channel 150 km inland from Townsville in Queensland where it was growing on fresh sand bars.
In the Northern Territory, a tropical American riparian shrub, Mimosa pigra , arrived at Darwin in 1947, believed to have come via Indonesia. In 1952 it was noticed along the banks of the nearby Adelaide River. By 1983 it was present in masses along various rivers across about 250 km of the Northern Territory coastal region. It is a spiny, rank-growing shrub, up to 4 m tall, that competes effectively with the native floodplain vegetation. The seed is dispersed by both water and animals and remains viable for long periods. Control is currently being attempted with imported New World insects.
Comment
Stream banks have evidently been primary breeding grounds for a large roster of colonizing species. Colonization by riparian species is commonly tightly integrated with river flooding, which opens up new habitat patches, disperses floating seed, and leaves moist sediment for seedling establishment. Vogl (1980) made the same point for trees growing along usually dry washes in the Mojave Desert of California. Flash floods destroy old populations and at the same time create suitable conditions for germination and establishment of new individuals. Some woody legumes, including the smoke tree, Dalea spinosa , require stream abrasion for seed germination.
For riparian species capable of perpetual micromigration, a drainage basin offers a relatively continuous and permanently available territory. However, riparian species are not usually confined to single drainage basins. In addition to being dispersed by water, they are evidently widely dispersed by wind or birds. Numerous cases of naturalization following even longer range human introductions show that species had not all filled their potential areas. Examples could be multiplied for other regions than those mentioned above. An extreme case occurs in New Zealand; according to Dansereau (1964), New Zealand stream banks have been so completely taken over by exotic plants that it is now unclear whether they have displaced native species or invaded an empty niche. The invaders are mostly European species of Salix, Alnus, Ulex , etc., but Lupinus arboreus from the California coast has joined in.
Debris Avalanches And Earthflows
White Mountains, New Hampshire
(Flaccus 1959)
Over 500 debris avalanches and earthflows were located on aerial photographs of the White Mountains. Vegetation was sampled across seven of these of known ages, ranging from 9 to 72 years; most were initiated by intense summer rains. Substrates were heterogeneous, including talus, glacial till, and bedrock. In general, where the slide left areas of stripped bedrock or rock debris, almost no revegetation had taken place. Areas where organic matter and soil were mixed into the displaced material were rapidly revegetated. On a 9-year-old slide, colonists of such areas included native meadow herbs, Achillea millefolium and Epilobium spp.; pioneer trees, Populus tremuloides, Betula lutea , and B. papyrifera ; and a Eurasian weed, Rumex acetosella .
On a 19-year-old slide, the pioneer herbs were almost gone in a dense thicket of aspen and birch. Picea rubens and Abies balsamea , the general dominants of adjacent mature forests, were rare on younger slides but were becoming dominant on a 72-year-old slide; there, shade-tolerant forest herbs were beginning to appear.
Gothic Mountain, Colorado
(Langenheim 1956)
In the spring of 1923, after a winter with twice the normal precipitation, saturated old morainal mantle slid off a steep slope taking some of the underlying friable shale with it. The slide extended from about 3,000 m down to about 2,750 m elevation.
A study 24 years later found the adjacent vegetation to be a sagebrush steppe, grazed by livestock; dominants were Artemisia tridentata and Festuca thurberi with groves of Populus tremuloides . On most of the slide, the dominant was Chaenactis alpina , a perennial herb that ranges widely through the high mountains of western North America on rock outcrops and talus. Other native pioneer herbs included Senecio atratus and Epilobium angustifolium. Populus tremuloides had colonized the slide but was not yet abundant. Eurasian weeds present on both the breakaway headslope and the debris toe included Poa pratensis, Sitanion hystrix, Bromus inermis , and Taraxacum officinale .
Valdivian Andes, Chile
(Veblen and Ashton 1978; Veblen et al. 1981)
In 1960, a great earthquake (8.75 on the Richter scale) in southern Chile set off thousands of dry debris avalanches, landslides, and mudflows. Aerial photographs showed that over 25,000 hectares of land slid, much of it on steep slopes blanketed with deep, water-soaked volcanic ash. The event was not unique; since the Spanish conquest, the region has had 7 comparable great earthquakes and 40 other strong ones. Volcanic activity is also frequent. The regional forests are typically even aged stands dating from slides or ash falls. Some older stands have been invaded by shade-tolerant, late successional species, but even where there has been no logging or clearing, equilibrium steady-state forests are perhaps not found anywhere. Various hardwood trees, mainly Nothofagus spp. and Eucryphia cordifolia , both genera having wind-dispersed seeds, dominate the forests of all ages.
In 1975–1976, vegetation on surfaces exposed by the 1960 slides and adjacent older surfaces was studied. In general, colonizing species were shared
between exposed ash and rock slides, although colonization of unstable ash was slower than on stable ash and rock slopes. Seedlings of the dominant trees in the surrounding forests had evidently begun colonizing within 4 to 5 years of the earthquake. On loose ash, the Nothofagus seedlings invaded gradually from the periphery, perhaps not because of dispersal limits but because of better substrate conditions, including available soil fungi that form symbiotic ectomycorrhizae on the roots.
Shade-tolerant shrub species in the adjacent older forests were rare or absent on the new slides. Some have heavy seeds, such as Persea lingue , but others are widely dispersed, and their failure to establish must be attributed to habitat problems. The common shrub colonists were mostly pioneer bird-dispersed species of forest edges, clearings, and burns, for example, Fuchsia magellanica, Coriaria ruscifolia , and Buddleia globosa . The native bamboos, Chusquea spp., which are extremely abundant in older secondary vegetation of the region, were missing from the slide surfaces. They flower and produce seed only at intervals of decades and vegetative spread is slow.
A native forest margin herb, Gunnera chilensis , colonized rock slides much more successfully than ash slides. It is a perennial with huge leaves and symbiotic nitrogen-fixing blue-green algae within its tissues; its succulent red fruits are probably attractive to birds. Most herb colonists on the slides are weedy Eurasian species, annuals and perennials, mostly wind dispersed, that probably seeded in from artificially disturbed habitats, for example, Poa pratensis, Holcus lanatus, Rumex acetosella, Prunella vulgaris, Digitalis purpurea , and Hypochoeris radicata .
Comment
Landslides are a normal feature of the earth's tectonically and volcanically active mountain systems. Innumerable examples could be cited; for example, an estimated 45,000 hectares of Darien Province, Panama, were denuded by earthquake-triggered landslides in 1976 (Garwood et al. 1979). Slides can be activated without earthquakes simply by saturation with heavy rains. Geomorphologists commonly use a Southeast Asian term, lahar , for slides with a high water content. Lahars are major contributors to the landscape of many regions, including northwestern North America (Franklin and Dyrness 1973). They are especially prevalent on volcanic ash but can occur on a variety of substrates.
Landslides do not have such discrete floras as the habitats previously discussed; they probably occur too erratically in space and time to support an unshared flora. However, it does not follow that this habitat has played an insignificant role in migration and evolution of pioneer plant species. Some generalist pioneer species may have always been shared between landslides and other disturbed habitats, all of which helped shape their adaptations
and distributions. For example, quaking aspen, Populus tremuloides , is one of the most cosmopolitan trees in North America; it pioneers in a wide variety of sites, including glacial outwash and volcanic cinder cones. A significant part of the species population lives on landslides in various mountain ranges. Aspen produces good seed crops every 4 to 5 years, releasing tremendous quantities of seed, buoyed by long silky hairs, that can be carried for miles by air currents. The ripe seed, which is produced in the spring, is nondormant and highly viable, although successful seedlings are infrequent. To survive, a seedling needs constantly moist, bare mineral soil. Once established, a single aspen can spread into an indefinitely large clonal grove by root suckering. Spread by suckering is stimulated by almost any kind of disturbance. A new and wet landslide provides ideal conditions for aspen seedling establishment, and continued instability aids clone expansion. If an aspen clone can escape being overwhelmed by competition from Picea or other later successional trees, within 20 years it will begin producing seed, which may colonize another landslide or other opening.
In the cases discussed above, except for the few species noted as bird dispersed, most of the pioneer colonists of landslides are capable of wide wind dispersal. This applies to both natives and introduced weedy species. However, the bulk of the populations of the latter are in artificially disturbed habitats, and landslides may be relatively insignificant as migrational pathways.
Recently Deglaciated Land
Mount Robson, British Columbia
(Tisdale et al. 1963)
Valley glaciers on Mount Robson, the highest peak in the Canadian Rockies, have been generally receding since 1780, with several halts during which recessional moraines were deposited. In 1963, vegetation was studied on three recessional moraines that had been abandoned by ice 50, 73, and 160 years before. The valley in which the moraines lie is obviously within the seed shadow of a dense, old Picea engelmannii forest growing on the unglaciated slopes above the glacial trim line.
In the moraine available for colonization for 50 years, plant colonization was confined to well-drained gravel and sand ridges. These had sparse, low-growing perennials, mainly Dryas drummondii, D. octopetala , and Hedysarum mackenzii , all three of which have root nodules with nitrogen-fixing bacteria. Rarer shrubs were Arctostaphylos alpina and Salix spp. The entire
roster are delegates, not from the adjacent spruce forest, but from high terrain above timberline. Dryas and Salix are wind dispersed; the others are dispersed by birds or streams.
Ridges exposed for 73 years still had the same species roster and a few Picea seedlings. On ridges exposed 160 years, Arctostaphylos was much denser, and the Picea tree canopy covered about 20% of the area. Wet swales between the ridges still had almost no vegetation.
Glacier Bay, Alaska
(Cooper 1939; Crocker and Major 1955; Lawrence 1958; Lawrence et al. 1967; Mirsky 1966)
Valley glaciers here have retreated nearly 100 km during the last 200 years, but most of the lost area has been invaded by the sea to form fjords. Revegetation of areas above sea level has been followed with the aid of maps and photographs dating back to 1886.
On the youngest moraines, which have been-ice free for 16 years, the first seed plant colonists, along with mosses and Equisetum , were Dryas drummondii, Salix arctica , and Epilobium latifolium . All of these are wind dispersed, presumably from above timberline; none are members of the nearby mature spruce forest, here dominated by Picea sitchensis . On slightly older moraines, these were joined by other Salix spp., Populus trichocarpa , and Alnus crispa , the last of which has root nodules with nitrogen-fixing bacteria, like Dryas .
The raw moraines had a high pH due to a high marble content, but as soon as plant colonization began, the acidity of the surface soil increased rapidly along with organic carbon and nitrogen compounds. During 20 to 40 years of dominance, Dryas increased the nitrogen content of the soil and plant system from about 35 kg/ha to about 400 kg/ha. Populus trichocarpa grew best after establishment of a Dryas cover. The carbon and nitrogen content of the soil increased more rapidly after arrival of Alnus and peaked after about 100 years under alder.
Picea sitchensis seedlings began to invade the alder thickets early but remained rare and stunted, the oldest trees in a given site being all very small compared to some younger spruce on the same site. From about 65 to 170 years after ice retreat, the spruce increased in growth rate and gradually replaced the alder entirely. After that, the nitrogen content of the soil declined. About 200 years after the area was free of ice, the spruce forest was invaded by hemlocks, Tsuga heterophylla and T. mertensiana . Occasional seedlings of these species had appeared much earlier but were suppressed. Like the first pioneers, all the later arriving trees are wind dispersed; it is inconceivable
that the sequence of immigration was controlled by differences in dispersal.
Poorly drained areas had a different history. The only common tree colonist was Pinus contorta . Eventually these areas became blanketed with wet, acid Sphagnum , which formed an expanding peat bog at the expense of conifer forest.
A sequence differing only in minor details was reported from moraines left by retreat of the Juneau Ice Field in Alaska (Crocker and Dickson 1957).
Rhone and Aletsch Glaciers, Switzerland
(Ludi 1958)
Valley glaciers here have been in general retreat since the mid-nineteenth century. The deglaciated valley floors lie below slopes forested with Picea abies , with Larix decidua higher up. Intensive study of areas that had been free of ice only 10 years or less showed that the first pioneers were alpine herbs that grow in meadows above timberline and along mountain streams, such as Saxifraga aizoides , a circumboreal arctic-alpine species. Some of these species were only present briefly and may have been derived from seed that had landed on the glacier and was deposited by meltwater.
About 10 years after ice retreat, some seedlings of woody species were present, for example, Alnus incana, A. viridis, Betula pendula , and Salix spp. By 25 years after exposure of the ground, Alnus thickets were established and producing seed; a few stunted individuals of Picea and Larix were present.
Laguna San Rafael, Chile
(Heusser 1964; Lawrence and Lawrence 1959)
By 1882, a lobe of the huge North Patagonian Ice Field that had reached tidewater in Laguna San Rafael began a slow recession. From 1910 to 1935 recession was rapid. Between 1940 and 1958, the edge remained nearly stationary, with a slight advance that overran some young forest just before the vegetation was studied in 1959. A total of 10 recessional moraines left between 1882 and 1940 were given approximate dates from historical records, maps, and tree rings.
Unglaciated mountain slopes above the glaciated valley bear a cool temperate rainforest dominated by hardwoods, Nothofagus spp., Weinmannia trichosperma, Drimys winteri , and a conifer, Podocarpus nubigens. Chusquea quila forms bamboo thickets where the forests have been cut or burned.
The youngest deglaciated surfaces available for study had been available for colonization about 25 years. Better drained moraines and outwash areas on those surfaces had been colonized by the gigantic herb, Gunnera chilensis , and its smaller mat-forming relative, G. magellanica ; both have nitrogen-fixing blue-green algae in their leaf axils. Other pioneers included tree ferns and flowering shrubs, such as various species of Empetrum, Pernettya , and Baccharis . Also, a few seedlings of rain forest trees, such as Nothofagus and Drimys , and a forest margin shrub, Fuchsia magellanica , had appeared but no Chusquea .
Better drained areas available for 50 to 75 years were covered with young Nothofagus and Drimys trees; the pioneer species were dying out and seedlings of Weinmannia and Podocarpus were establishing. Poorly drained areas on all these surfaces had grassy marshes and ponds with freshwater aquatics, such as Potamogeton and Myriophyllum spp.
Comment
In the cases cited here, freshly deglaciated terrain clearly presents edaphic problems that bar immediate immigration by species from adjacent mature forests. On better drained moraines, these barriers are evidently gradually relaxed after roughly 50 years, evidently by changes in soil under pioneer vegetation; nitrogen fixation by symbiotic bacteria may be crucial. Invasion of poorly drained areas is even slower.
Modern deglaciated terrain is too restricted in time and space to have a special flora. The pioneer colonists have a variety of primary habitats, including alpine tundra, mountain streambanks, and scree. All are adapted for fairly long-range dispersal, many by wind.
Emerging Nonvolcanic Islands
Baltic, Finland
(Barlund 1940; Luther 1961)
The Baltic Sea is constantly gaining new islands as isostatic rebound of the earth's surface, relieved of the weight of the continental glacier, still proceeds. The floras of 22 tiny islets off the southwest coast of Finland have been censused repeatedly, in most cases at least three times over a period of
more than 50 years. The islets are mostly bare granite and gneiss scoured by glacial ice. The largest is about 4 ha in area and about 10 m high. At the calculated rate of emergence of about 45 cm per 100 years, the oldest island has been exposed for about 3,000 years and the youngest about 150 years.
A total of 174 species of seed plants and 7 species of ferns were recorded on the 22 islets. One island had no flora in three censuses spanning over 50 years; it was not the smallest or lowest of the islands but one of the most isolated and exposed. Although about 2 m high and about 400 years old, it was still washed over by storm waves. Numbers of species in the floras of the other islands ranged from 1 to 108 in a given census. Ten of the islands had 1 to 15 species at their first census, and only five of these had more than 15 species at their last census. These low counts were found on islets where plant colonization was restricted to crevices in otherwise bare rock. The commonest species in these crevices were Allium schoenoprasum, Juncus gerardi, Festuca rubra, Puccinellia retroflexa, Agrostis stolonifera, Rumex crispus, Sagina procumbens, Cochlearia danica, Sedum acre, and Matricaria maritima . These are perennial herbs that are widespread in Europe, mainly in salt marshes and rocky seashores; some also grow in open inland habitats. None have obvious mechanisms for long-range dispersal, but they are believed to be carried by birds and flotation.
The other 11 islands had over 20 species at each census. Usually these included all the species noted above in rock crevices. Additional species were found in two other habitat types: pocket beaches and interior flats and hollows beyond reach of the waves. Beaches were colonized intermittently by a few common sea-dispersed species: the annual Cakile maritima ; the perennial vine, Lathyrus maritimus ; and the perennial grass, Elymus arenarius . These colonists did not always persist and may have been recruited repeatedly from the mainland.
Sheltered interior flats and hollows had a richer flora, both wind and bird dispersed. Hollows with freshwater pools had Typha latifolia, Lemna minor, and Triglochin palustre . Fringing the pools were marshy meadows, mossy bogs, and heaths with a rich mixture of sedges, rushes, and grasses, including Poa pratensis and Phragmites australis , and dicot herbs, including Galium palustre, Epilobium plaustre, and Cornus suecica . Shrubs included Juniperus communis and a variety of heaths and their relatives, particularly species of Calluna, Empetrum, and Vaccinium . Tree species mainly colonized the interiors of the largest islands. The only tree present during the first census was Sorbus aucuparia . By the later censuses, Pinus silvestris, Picea abies, Populus tremula, Betula pubescens, B. verrucosa, Salix aurita, and Alnus glutinosa were present.
Successive censuses found many changes in the floras of most islets. For example, one medium-sized islet was censused five times over a 62-year
period; it initially had 43 species of which 34 were found in all succeeding censuses. The number of species present increased with each succeeding census to 89 in the final count. A total of 103 species were found in at least one census, of which 25 disappeared at least temporarily in later censuses. A similar pattern was found on the other islets: almost every census showed more species than the previous one, but often a few species had died out. These ephemeral colonies often consisted of a single or few individuals, often nonflowering. The islets are probably continuously inoculated with seed of beach plants and inland pioneers produced by mainland populations upstream and upwind.
At the last census there was an outburst of seedlings of weeds and crops on islets used as nesting grounds by gulls. Shortly before, the gulls of south Finland had shifted their foraging behavior by increased feeding in agricultural fields and rubbish dumps. Luther (1961) found regurgitated gull pellets responsible for much seed dispersal. Seabirds evidently were also responsible for a few cases of species extinction. One islet originally had a stand of shrubs, including Juniperus, Empetrum, and Vaccinium, which died out completely after a buildup of nesting birds. Extinction of several herb species was attributed to an exceptional buildup of voles.
Plants on these rocky, windswept islands might be expected to be highly vulnerable to drought. Two exceptionally dry years occurred between the last two censuses; August in one of those years was the driest since records had been kept in southern Finland. On seven islets, several species present in small numbers before the drought apparently died out. Dieback of other species was heavy, but recovery by sprouting and seedlings was rapid in most cases.
The steady immigration of new species during the study period is not easily explained. For example, why should the highest island, which has existed for about 3,000 years, have had 33 species at the first census and 58 species 50 years later? On some of the other islands, the increases also seem too abrupt to be due to fortuitous dispersal of species that had not happened to arrive before. Uplift of about 25 cm during the study period would have increased the islet areas only slightly. In some islets, uplift might have raised surfaces above a threshold, beyond reach of sea ice and storm waves. Once freshwater pools are beyond reach of the sea, a slow process of autogenic succession could begin. As peat and turf develop and expand over surrounding rock surfaces, colonization by heath and tree species might begin. Also, there may be a positive feedback between complexity of an islet's flora and vegetation and the amount of bird traffic visiting the islet.
The islets discussed above are mainly bare rock. The processes of colonization and succession evidently proceed faster on nearby islets composed of loose morainic material. For example, in the Ahvenanmaa Islands, some islets
that emerged only 250 years ago already have thickets of Alnus glutinosa and Hippophae rhamnoides .
Gulf Of Bothnia, Finland
(Vartianen 1980)
Isostatic rebound here is even faster, about 75 cm per 100 years, creating shoals of new islands composed of glacial debris that has been reworked by the sea. Plant colonization has been studied by two complementary methods.
Time-lapse vegetation maps were made for four relatively large, old islands that emerged from the sea about 1,000 years ago. Maps in the national archives of the Surveyor General were used to plot vegetation in the years 1770, 1880, 1930, and 1958. Aerial photographs were used to plot vegetation in 1977. All the maps were reduced to the same scale, and they show distributions of sand beaches and dunes, boulder shores and ridges, sedge and grass meadows, swamps, lagoons, fens, heath scrub, deciduous woodland, and pine woods.
The map sequences show fascinating changes as the four islands rose and expanded. The changes may have been modified by sheep grazing, which continued on the islands until the 1940s. The A.D. 1700 maps, made when the islands were more than 700 years old, show bare sand and boulder areas, meadows, and heaths, but no woods. By 1880, the same types, greatly expanded in area, still covered the islands. By 1910, there were small patches of deciduous woodland and swamps in the interiors of the islands, while the meadow and heath areas were displaced toward the expanding peripheries. By 1930, deciduous woodland had expanded on some islands but had changed to a swamp on another; there were small areas of pine woods. The greatest change in vegetation came between 1930 and 1958 when the central deciduous woodlands expanded greatly into former heath and meadow areas; there was also some expansion of pine woods. Perhaps this was partly because the islands had passed a critical threshold of size or age before woodland could develop, and perhaps it was partly because of sheep removal. Between 1958 and 1977, there was less change.
Another study method used belt transects of 150 islands in the same archipelago to characterize species patterns in the modern vegetation. The data have no time depth except as the height above sea level indicates the time available for plant colonization since emergence. Most of the islands are ungrazed, but some have been visited by sealers and fishermen. Nearly 300 species were found. Their patterns show a rough zonation correlated with elevation and thus with age of the surface since emergence. Sea ice pushed ashore by the wind affects the lower margins of vegetation.
Lake Hjälmaren, Sweden
(Birger 1907)
In 1882–1886, this central Swedish lake was partly drained, its level being lowered about 2 m to reclaim 15,000 ha of farmland. A large residual lake remained in which 29 new islets had emerged; although none were more than 2 m high, 26 of them were no longer washed over by the gentle waves of the lake. Most of the new islets were separated by at least a few kilometers of water from older land surfaces. The islets were partly cracked rock surfaces and partly morainic material.
Plant colonists were censused after 4, 10, and 22 years. The first census found 113 plant species, mainly freshwater marsh plants and common annual agricultural weeds, but also found Betula verrucosa, Populus tremula, and Alnus glutinosa . Possible dispersal agents include wind, birds, water currents, and floating ice. It is unknown which species had dormant seed on the lake floor before drainage.
A total of 260 plant species were found in the three censuses combined, many only once. Most islands peaked in species number 10 years after emergence. The pioneer hardwood trees were then joined on some islands by Pinus silvestris and Picea excelsa . By 22 years after emergence, Quercus robur and Tilia europea had appeared; by then some of the pioneers were being crowded out by the developing tree cover. For example, the marsh annual Bidens tripartitus was present on 14 islands at the first census, all 26 islands at the second census, and 10 islands at the third.
Comment
This set of cases shows how difficult it is to generalize about colonization of new islands, even within a relatively restricted and climatically homogeneous region. Some islets had a rich flora, including several tree species, within 4 years of emergence. Another islet of similar size had no plants 400 years after emergence. One archipelago began acquiring trees 900 years after emergence; another was nearly 3,000 years old before it began acquiring trees. Some islands reached a peak in number of plant species 10 years after emergence, and then species numbers declined as pioneers were eliminated by competition. Other islands that were thousands of years old doubled their species numbers during 50 years.
It is impossible to explain such differences by any model based simply on quantitative measures of islands, for example, area, height, distance from seed sources, and age. Habitat quality must be included as a crucial variable. In these cases, the most obvious habitat variables are exposure and substrate.
The most rapidly colonized islands were in a lake with low-energy waves; their substrate included loose glacial till and water-borne sediment. The islands with the slowest colonization rates were exposed to sea waves and brackish spray; their surfaces were mainly rock. Islands with intermediate colonization rates had a complex of different habitats. Habitat complexity may be partly a function of island area and height, but the relationship is not linear. Habitat complexity may increase abruptly when size and height thresholds are passed. Once interior areas are beyond reach of waves and spray, a new flora can colonize and initiate successive changes that create opportunities for additional immigrants.
This set of cases all involves islands that are rather close to seed sources; the islands are probably within the normal seed shadow of far more species than have immigrated. In other words, screening of the colonizing species was generally by environmental selection rather than by dispersal barriers. However, dispersal has not necessarily been constant; for example, a change in seagull foraging patterns evidently led to an outburst of ephemeral crop and weed seedlings on gull nesting lands. Extinction of these unsuccessful immigrants was evidently due to lack of suitable habitats. Also, a few cases of temporarily successful immigrants becoming extinct are attributable to habitat change, for example, pioneer species being eliminated by succession and marginal populations being killed during extreme drought.
Fresh Lava And Pyroclastic Deposits
Paricutin, Mexico
(Beaman 1961; Eggler 1963; Rees 1979)
The town of Paricutin, Michoacan, was built on the slopes of a great Pleistocene volcano, which had been dormant long enough for even the steeper cinder slopes and lava benches to become heavily wooded. There were forests of fir, Abies religiosa, at high elevations, woodlands of Pinus and Quercus species at intermediate levels, and complex broadleaf subtropical forests in the deep barrancas. In the pine—oak zone, gentler slopes with good soil had mostly been cleared for milpas.
The new volcano began erupting in 1943 and continued to be active until 1951, eventually engulfing the town. Although a relatively small flank volcano, it emitted over 2 billion tons of ash and other pyroclasts and over 1 billion tons of lava, mostly of the blocky aa type. Lava flows and cinder cones wiped out all vegetation on about 2,500 ha; ash falls caused partial
kill on about 35,000 ha. Pines and oaks survived ash falls better than the higher elevation firs and the lower elevation subtropical trees. In milpas covered with shallow ash, only a few weeds survived, notably Cynodon dactylon , an African grass now ubiquitous in weedy vegetation of warm regions. Attention here will be focused on areas where revegetation was by colonization, not regrowth.
Colonization on lava flows proceeded faster than on cinder cones or ash. By 1960, lava flows had abundant blue-green algae, lichens, mosses, and ferns; crevices and pockets had scattered individuals of over 30 seed plant species. By 1978, young trees of three species were common on lava: Pinus leiophylla, Clethra mexicana, and Buddleia cordata . Early colonists of cinder cones included a few ferns; native shrub species, such as Phytolacca icosandra, Wigandia kunthii ; and cosmopolitan weeds, such as Digitaria sanguinalis, Conyza canadensis, and Sonchus oleraceus . Areas mantled with deep ash remained barren until the ash was enriched with water-transported soil or organic debris or where erosion exposed underlying soil. Such areas were colonized by shrubs and herbs native to rocky slopes and ravine banks; most were composites, for example, Baccharis, Eupatorium, and Senecio spp., or legumes, for example, Dalea, Desmodium, and Crotalaria spp. One was a Ceanothus species, which, although not a legume, probably has nitrogen-fixing root bacteria.
A few of the colonizing species are adapted for bird dispersal, including Pinus, Phytolacca, and Gaultheria , but most are wind dispersed. Screening of colonizing species from the rich regional flora is, however, obviously mainly by habitat conditions rather than dispersal limits.
Irazu, Costa Rica
(Parsons 1972)
This huge ancient volcano resumed activity in 1964–1965. Ash falls on the slopes below killed many oaks and other native trees and buried the grasses and other herbs in areas cleared for dairy pastures. After the eruption was over, the ash falls were rapidly invaded by Kikuyu grass, Pennisetum clandestinum , native to the volcanic highlands of Kenya. Kikuyu grass had been introduced to Costa Rica about 1928 but was generally considered a pest until the eruption. Because of its vigorous colonization of the ash falls, it has become the preferred grass for dairy pasture at elevations between 1,800 and 3,000 m on Irazu.
In 1976, I found that the new cinder cone at about 3,200 m elevation was still mostly devoid of vegetation. There were patches of Rumex acetosella, R. obtusifolia, and Melilotus alba , Eurasian annuals that are now cosmopolitan
weeds in temperate climates. There were also patches of the native Lupinus aschenbornii and Coriaria ruscifolia. Coriaria ruscifolia is a strikingly unusual shrub with a highly disjunct range in the American Cordilleran system at high elevations and in the southern hemisphere at high latitudes. In the Cordillera it ranges from the western Sierra Madre of Mexico down to Chile in a variety of open habitats, including ravine banks, landslides, and burns. The same species is native to New Zealand, where it was a prominent colonist of the Tarawara Volcano after a great eruption in 1886 (Cockayne 1927). The seeds are enclosed in a succulent fruit and are presumably bird dispersed. Although not a legume, Coriaria has symbiotic nitrogen-fixing bacteria.
Cameroons Mountain, Cameroon
(Keay 1959)
This colossal volcano towers more than 4,000 m above the shore of the Gulf of Guinea. An eruption in 1922 left a fresh lava flow 10 km long and over 1 km wide extending from 1,400 m elevation down to the sea. The adjacent vegetation was a tropical rainforest, the region being one of the wettest in the world. About 15 years later, the lower elevation parts of the lava flow had been colonized by various spore plants and over 40 species of flowering plants. Another study 29 years after the eruption, made by a different person in a slightly different place, produced an almost identical plant list, with only 5 additional species.
The dominant colonist was a tree in the coffee family, Hymenodictyon biafranum , common on open rocky sites in the West African mountains; its seeds are wind dispersed. The next most abundant was a dragon's blood tree, Harunga madagascariensis , with a wide, disjunct range extending to Madagascar and Mauritius. It grows in rocky banks, along streams, and in wet swales; it is evidently bird dispersed. Also bird dispersed were several species of Ficus and Psidium guayava . The figs are native; the guava was introduced from the American tropics as a planted fruit tree soon after 1500 A.D. and promptly escaped. Another common colonist on the Cameroon lava flow was kapok, Ceiba pentandra , one of the world's most enormous trees, also native to the American tropics where it is a pioneer in riparian and other open sites. Baker (1965b ) concluded that Ceiba arrived in West Africa in ancient times, probably by means of ocean-drifted fruits, and spread across Africa as a pioneer of riparian and other open sites. Its seeds are produced in huge quantities and are easily carried by the wind after a mature fruit dries and breaks open.
The lava flow had also been colonized by various herbs, both wind and bird dispersed, including Axonopus compressus, Paspalum conjugatum, Asystasia gangetica, Borreria ocymoides, and Emilia coccinia . These are mostly of doubtful nativity and are now common pantropical weeds.
Mauna Loa, Hawaii
(J. O. Juvik, personal communication 1977; Smathers and Mueller-Dombois 1974)
Kilauea is a still-active volcano at about 1,200 m elevation on the lower slopes of Mauna Loa. Kilauea has produced many prehistoric and historic lava flows, cinder cones, and ash beds. Under the tropical rainforest climate, a dense forest dominated by Metrosideros polymorpha has developed on some of the prehistoric lava, including the floor of a satellite crater, Kilauea Iki. In 1959, an eruption of Kilauea Iki buried this forest under a lava lake 100 m deep and 56 ha in area; the lava was mainly of the smooth, ropy pahoehoe type. The same eruption built a cinder cone covering 19 ha and scattered pumice over a larger area, in part of which the pumice was deep enough to cause total kill of the vegetation.
A study done 9 years after the eruption found that the lava lake had not yet cooled in the center, which steamed after every rain. Even where no longer heated from below, the dark lava was heated intensely by the sun, and water retention on the smooth pahoehoe was nil except in crevices. Around the totally bare, hot center was a wide concentric zone sparsely colonized by algae, mosses, and ferns. Although the lava lake lay downslope and downwind from an undamaged rainforest, less than 1% of the lava surface was colonized by seed plants. The species that had gone farthest onto the lava lake were three endemic woody sclerophylls present in the adjacent forest: Vaccinium reticulatum , which had colonies as far as about 100 m from the edge and about one-third of the way to the center, followed by Metrosideros polymorpha and Dubautia scabra . Behind them, edging onto the first few meters of the lava, were a motley group of soft-leaved perennials, mostly exotics. These included Lythrum maritimum , native to streambanks in the Peruvian coastal desert, which had somehow become naturalized in the mountains of Hawaii in the nineteenth century; Andropogon virginicus , native to savannas and marshes of eastern North America; and three recent garden escapes, Anemone japonica, Buddleia asiatica , and Fragaria vesca .
Moisture conditions on the loose pyroclastic deposits were not much better than on the lava. Water available to plants, defined as the difference between capacity at saturation and the permanent wilting point, was about 3% of the weight of the ash, less than in pure sand. By 9 years after the eruption, the cinder cone had 6 species of seed plants; other areas of total kill under deep pumice had 21 species. The most common species, both native and exotic, were the same as on the lava lake.
Beyond the total kill area, survival increased with decreasing depth of pumice. The Vaccinium, Metrosideros, and Dubautia resprouted even when badly damaged, and seedlings appeared in abundance; thus, retreat of the original species was trivial. The area of partial kill did, however, open a
temporary opportunity for colonization by exotics. About 40 exotic species were present there 9 years after the eruption, including all those found on the lava lake. Two of the most successful were an Asiatic raspberry, Rubus rosaefolius , and an American blackberry, R. penetrans . There were also many sedges and grasses, including Pennisetum clandestinum , Kikuyu grass. However, Kikuyu grass did not blanket pyroclastic deposits here as it did on Irazu (discussed above). On Kilauea Iki in 1977, I found it abundant only on roadsides.
In the same general region in 1977, Juvik conducted a field trip to examine older lava flows, including an 1855 flow on the northeastern side of Mauna Loa. This flow, largely of the blocky aa lava, extended from above 1,800 m elevation to below 1,000 m. The upper part is above the trade wind inversion, has only about 125 cm of rain per year, and was still extremely barren; the lava looked unweathered and very fresh. The middle level, about the same elevation as Kilauea Iki, has about 500 cm of rain per year, and it had a sparse vegetation dominated by the same species of Vaccinium, Metrosideros, and Dubautia that colonized the lava lake on Kilauea Iki. The lower level has about 750 cm of rain and had a similar but denser vegetation.
Metrosideros remains a dominant on the most ancient Hawaiian aa lava flows over a broad range of annual precipitation. This is a good example of the initial floristics model of succession, in which the later dominants are among the pioneer colonizers. Old Metrosideros stands are subject to a patchy dieback, but in a cyclical mortality and regeneration process that does not eliminate it from the vegetation (Mueller-Dombois 1980).
Mount Komagatake, Hokkaido, Japan
(Numata 1974)
In 1929, a violent eruption deposited pumice over a large area of Mount Komagatake from 600 m elevation down to 200 m. The theoretical climax dominant species of forests in the region is Fagus crenata , but the actual forests are mainly secondary, dominated by Quercus mongolica and species of Populus, Betula, Castanea, Prunus, and Sorbus . In the area of total kill, plant seedlings began appearing in 1932 in interstices between the pumice blocks. The colonizing species were mostly wind-dispersed pioneers common in disturbed habitats of the region. The first trees to invade were Salix, Betula, and Populus . The first herbs were widespread weeds: Miscanthus sinensis, Chenopodium album, Epilobium angustifolium, and Anaphalis margaritacea . By 1935, these were joined by other cosmopolitan weeds, including Poa annua and Rumex acetosella , and by a garden escape, Oenothera grandiflora . By 1942, the weedy herbs were being shaded out. By 1960, areas sheltered from the wind had a dense young forest of Populus, Betula , and Salix, with seedlings
of other natives, including Acer, Prunus, Hydrangea, and Rhus . By 1965, this forest had not changed in composition.
Volcanoes on Honshu, Japan
(Numata 1974)
Dated lava flows on several mountains on Honshu permit age and elevation comparisons. Mount Zao had a thick andesite flow about A.D. 750 in the subalpine zone, where Abies mariesii is the theoretical climax dominant species. At the time of study, saplings of this fir had just begun to colonize. The vegetation was still dominated by gnarled scrub of Pinus pentaphylla , with an undergrowth of Empetrum nigrum and other dwarf alpine plants that had moved downslope from their usual zone.
Mount Fuji had a basalt flow in A.D. 864 in a zone where Fagus crenata is the climax dominant. The lava is only slightly weathered, but a recent study found a dense growth of mixed conifers belonging to six genera. Pioneer hardwoods had already begun to colonize, particularly Quercus mongolica and Betula spp.
Mount Asama had an andesite flow in 1783 that extended from the subalpine zone, where the theoretical climax is a conifer forest dominated by Abies mariesii and Tsuga diversifolia , down into the Fagus crenata –dominated zone; at the same time, deep pumice was deposited in the lower zone. A recent study found the subalpine lava flow had been colonized only by lichens, mosses, and a few alpine seed plants, mainly ericads. The lower lava also remained barren of seed plants other than alpine ericads, except that sheltered hollows had scrubby thickets of Pinus, Larix, Alnus, and Betula . Pumice deposits in the lower zone had a dense, young pine forest, with an understory of Quercus, Prunus, and other hardwood seedlings.
Krakatau, Indonesia
(Borssum Waalkes 1960; Docters van Leeuwen 1936; Richards 1964; Whittaker et al. 1984)
The famous eruption of 1883 transformed the old Krakatau Islands, which had been covered with dense forests, into a wasteland of hot pumice and ash. The three new islands are all rugged, the highest peak being over 800 m elevation. Revegetation has been studied by repeated scientific expeditions.
The rapid immigration of sea-dispersed beach species and the assemblage of a rich coastal flora was noted in the section on Tropical Sea Beaches.
Evidently the distances from seed sources were insignificant for this flora, and the environmental obstacles to colonization of fresh pyroclastic beaches were significant for only a few of the member species.
For the inland flora, which is the object of attention here, both dispersal and environmental obstacles have evidently been effective in screening colonizing species. The overseas journey of about 80 km from Sumatra or about 45 km from Java has probably been accomplished by only a fraction of the bird-, bat-, and wind-dispersed floras of those islands.
An 1886 expedition 3 years after the eruption found the only plant life on most of the island surfaces to be blue-green algae and ferns, of which 11 species were reported. There were only 15 seed plant species, mostly sea-dispersed beach plants, and a few wind-dispersed grasses and composites. In the absence of competition, some of the vines normally confined to the beach outpost zone, such as Canavalia maritima and Ipomoea pes-caprae , were moving inland.
By 1897, the inland invasions of beach species were beginning to be contained by competition from inland pioneers that had not been present in 1886. In particular, two coarse perennial grasses, Imperata cylindrica and Saccharum spontaneum , had begun forming savannas over the lower hills and valleys. Both are extremely common and widespread in both naturally and artificially disturbed sites in southeastern Asia and the East Indies. They soon displaced the pioneer ferns and inland incursions of beach plants.
In turn, the savanna grasses were eventually displaced, except on special sites such as steep ridges, by development of a tree canopy. The first trees to arrive were Macaranga tanarius and three species of Ficus . All of these are common on riverbanks and in secondary woodlands in the East Indies, and all are probably dispersed by birds and fruit bats. By 1908, these four species were widespread on Krakatau and fruiting heavily. By then they had been joined by Neonauclea calycina , a secondary rainforest tree from Java which is wind dispersed, and also by Cyrtandra sulcata and Leucosyke capitellata , both understory species in mountain forests of Java and both bird dispersed. Two other bird-dispersed species that are American natives, Carica papaya and Lantana camara , also arrived in Krakatau by 1908; both had escaped from cultivation in the East Indies long ago.
On Krakatau a positive feedback evidently developed between increasing attractiveness of the vegetation to fruit-eating birds and increasing introduction of bird-dispersed species. The earliest plant colonists had been entirely sea and wind dispersed; by 1900, only six bird-dispersed species were present. Since then, the number of bird- and bat-dispersed species has increased over ten-fold, while the number of sea- and wind-dispersed species has increased comparatively slowly.
The first reports of weeds believed to be introduced by human agency were in 1919. From 1916 to 1921, Krakatau was inhabited by a small group of people engaged in pumice extraction. During that time, 17 weedy species
were found on the island for the first time, mainly near the boat landing and around the settlement. An example is Eleutheranthera ruderalis , an American native that had been introduced to the East Indies via botanical gardens; it had escaped in the nineteenth century to become a common weed in Java and Sumatra. Only 5 of these 17 species remained in 1929, the rest probably having been eliminated by competition.
The peak rate of immigration of new species evidently was reached in the 1920s when the developing forest canopy provided suitable habitats for more epiphytes and for species requiring shade and humus.
By 1951, behind the narrow fringe of beach vegetation, forests dominated monotonously by Neonauclea covered almost the entire main island up to 750 m elevation. Macaranga and various Ficus species were subdominants. On the summit above 750 m, Neonauclea and Macaranga were present but did not form a closed canopy; there was a dense shrub layer and patches of Sacharum grassland.
In 1979, a Hull University expedition found that the coastal vegetation had changed little in 60 years, except that Casuarina had lost most of its former area to broadleaf coastal trees. A few more inland species had arrived, such as Dysoxylum caulostachyum , a tree characteristic of primary lowland rainforest in Java, presumably bird or bat dispersed. The rate of immigration appeared to be slowing. Over the preceding 50 years, known additions to the flora averaged about 25 species per decade, while apparent subtractions averaged about 20 species per decade, giving a net observed increase of 5 species per decade. The successive surveys, however, were not equally intensive nor did they sample the same areas, so it is not known what proportion of the apparent species turnover is real.
The cumulative total of all seed plant species observed on Krakatau is slightly more than 300. Even if all these were still present, the flora is not nearly as rich as in comparable terrain in Java or Sumatra.
Surtsey, Iceland
(Fridriksson 1975)
The new island of Surtsey appeared above the sea in 1963. When the eruption was over in 1964, the island had an area of a few square kilometers, about half ash and half ropy pahoehoe lava. Only scientific research workers have been allowed to visit the island; all persons and equipment have been inspected to prevent accidental species introductions. However, some tubs filled with fresh water have encouraged bird visits.
The new island is less than 6 km from an old islet and not much farther from Iceland proper, which has a flora of several hundred seed plant species. Researchers on Surtsey have recorded arrival of seed in abundance, mostly
of species common in Iceland. Some is wind dispersed, for example, Eriophorum, Taraxacum , and Senecio . Seedlings have been found in sea gull droppings and carcasses. Snow buntings from Britain, stopping over on their spring northward migrations, are credited with bringing seeds of marsh plants, such as Polygonum persicaria and Carex nigra . None of these were successful colonists. The only seed plants that have survived to maturity on Surtsey are Cakile arctica and Elymus arenaria , both sea-dispersed beach plants; they may be repeated immigrants from Iceland rather than self-reproducing local populations. No inland seed plant species were established on Surtsey, only blue-green algae, mosses, and other lower plants.
Comment
Fresh volcanic deposits are too erratic in occurrence in space and time to support a discrete flora. Pioneer colonists are a hodgepodge of delegates from various habitats. If pyroclastic deposits are on a seashore, colonists arrive by sea dispersal from ordinary beaches. Inland pioneers are mostly wind or bird dispersed and come from rock outcrops, ravine banks, landslides, burns, and other disturbed habitats; some are weeds and escapes from cultivation.
Because dispersibility and ability to establish in disturbed habitats tend to be correlated, it is hard to sort out their relative importance in determining the roster of immigrants. In the cases of Krakatau and Surtsey, it is unclear how much effect the ocean barrier had on colonization patterns. In all cases, it is obvious that colonization by many species was not barred by failure of seed to arrive but rather by habitat. Fresh lava and pumice are clearly an impossible substrate even for some pioneer species. Lack of lime in pyroclastic beaches evidently prevents establishment of certain beach plants. A more general barrier is evidently lack of available nitrogen; prominent among early colonizers are both legumes and other plants with symbiotic nitrogen-fixing bacteria, such as Casuarina, Ceanothus, Coriaria , and possibly Saccharum .
The case histories generally suggest a relay floristics succession, with the pioneers modifying soil and microclimate until replaced by later colonists. However, the process is slow, even in humid tropical lowlands, and the available histories mostly show early stages.
The Hawaiian succession is anomalous, approaching an initial floristics model. The pioneers on fresh volcanic deposits include Metrosideros polymorpha , the ultimate dominant in old rainforests. For a rainforest tree, Metrosideros is also exceptional in its capacity for long-range dispersal. Its natural distribution extended from New Zealand and Australia through much of Melanesia and Polynesia. In the center of its range it is mainly a forest tree and is important as a pioneer on lava flows on the periphery of its range in New Zealand and Hawaii.