Appendix 1—
Mammalian Systematics
Brief descriptions of mammalian components of the original Bug Creek assemblages were given by Sloan and Van Valen (1965), Van Valen and Sloan (1965), and Van Valen (1978). Except for the few studies noted here, detailed systematic treatments have yet to appear. Lillegraven (1969) provided additional description of Procerberus . Archibald (1982) described some of the mammals found in the Bug Creek assemblages, but the focus of his systematic work was on mammals from Garfield County. Novacek and Clemens (1977) studied the UCMP sample of Mesodma from the Bug Creek Anthills locality and argued that only a single species was discernable (contra Sloan and Van Valen, 1965). Luo (1989, 1991) studied the "condylarth" component of the UCMP sample from Bug Creek Anthills and argued that three species are present rather than the single species, Protungulatum donnae , described by Sloan and Van Valen (1965). Lupton et al. (1980) briefly described two partial dentaries of Protungulatum gorgun from Chris' Bonebed, a Milwaukee Public Museum locality in the upper Hell Creek Formation near Harbicht Hill (Figure 1c).
The original Bug Creek faunas (Bug Creek Anthills, Bug Creek West, Harbicht Hill) formed the basis for the "Bugcreekian" NALMA (Sloan, 1987; Archibald, 1987b), but are now recognized as comprising the initial interval zone (Pu0) of the Puercan Age (Archibald and Lofgren, 1990).
Lancian mammals whose presence in Puercan faunas (Pu0 or Pu1 interval zones) in eastern Montana may be entirely caused by reworking are: Meniscoessus robustus, Essonodon browni, Cimolodon nitidus, Pediomys hatcheri, P. krejcii, P. florencae, P. elegans, Pediomys sp. indet., Alphadon "wilsoni," A. rhaister, A. marshi, Alphadon sp. indet., Glasbius twitchelli, Didelphodon vorax, Gypsonictops illuminatus , and Batodon tenuis . These questionable occurrences in Puercan faunas are denoted by an asterisk (*) in the listed distribution of each species. Cimexomys minor and Neoplagiaulax burgessi are Lancian mammals whose present in Puercan faunas apparently is not the result of reworking from Cretaceous sediments.
Lithostratigraphic units that yield both Puercan and Lancian mammalian local faunas are the Hell Creek Formation in McCone County, Montana, and the Frenchman Formation in Saskatchewan, Canada.
Class MAMMALIA Linnaeus, 1758
Subclass ALLOTHERIA Marsh, 1880
Order MULTITUBERCULATA Cope, 1884
Suborder PTILODONTOIDEA (Gregory and Simpson, 1926) Sloan and Van Valen, 1965
Family NEOPLAGIAULACIDAE Ameghino, 1890
Mesodma Jepsen, 1940
Mesodma sp.
Comments : A large number (n>2200) of isolated and associated teeth from fragmentary dentaries and maxillas referable to Mesodma were recovered from localities in the upper Hell Creek Formation at McGuire Creek sampled by screenwashing techniques. Distinction of species of Mesodma is a difficult taxonomic problem (Sloan and Van Valen, 1965; Novacek and Clemens, 1977; Archibald, 1982). Because of the large number of fossils and the difficulties inherent in species identification, a thorough study of the sample of Mesodma from McGuire Creek is beyond the scope of this study. Although it is uncertain if one or more species are present, all specimens are referred to Mesodma sp. at this time.
Family CIMOLODONTIDAE Marsh, 1889a
Cimolodon Marsh, 1889a
Cimolodon nitidus Marsh, 1889a
Cimolodon nitidus Marsh, 1889a, p. 84 (see Clemens, 1964, p. 56, for synonymies).
Holotype . YPM 11776, left /M1 (Marsh, 1889a, pl. II, figs. 5-8).
Type locality . Mammal locality no. 1 of Lull (1915), UCMP loc. V5003, Lance Formation, Wyoming.
Referred specimen . 1 M1/, UCMP 133406, from loc. V87072.
Locality . UCMP loc. V87072.
Distribution . St. Mary River Formation, Alberta (Edmontonian); Lance Formation, Wyoming, and Scollard Formation, Alberta (both Lancian); Frenchman Formation, Saskatchewan, and Hell Creek Formation, Montana (both Lancian and *Puercan); Ravenscrag Formation, Saskatchewan (*Puercan).
Revised diagnosis . Clemens, 1964, p. 56.
Description . The single tooth referable to Cimolodon nitidus is an M1/. The cusp formula (5:7:5) and size (L: 4.51, W: 2.57) of this specimen agree with Clemens' (1964) description of the species.
Suborder TAENIOLABIDOIDEA (Granger and Simpson, 1929)
Sloan and Van Valen, 1965
Family EUCOSMODONTIDAE (Jepsen, 1940)
Sloan and Van Valen, 1965
Subfamily EUCOSMODONTINAE Jepsen, 1940
Stygimys Sloan and Van Valen, 1965
Stygimys kuszmauli Sloan and Van Valen, 1965
Tables 12-13; Figures 13-19
Eucosmodon gratus Jepsen, 1930, p. 499-500.
Stygimys gratus Sloan and Van Valen, 1965, p. 224.
Holotype . UMVP 1478, left lower jaw fragment with I and alveoli for /P4 and /M1 (Sloan and Van Valen, 1965, part of fig. 4).
Type locality . Bug Creek Anthills, Hell Creek Formation, Montana.
Referred specimens . Three /P4's, 4 /M1's, 2 M1/'s, 3 /M2's, 7 M2/'s from loc. V87035. Six P4/'s, 4 /P4's, 8 /M1's, 9 M1/'s, 2 /M2's, 9 M2/'s from loc. V87037. Five P4/'s, 12 /M1's, 13 M1/'s, 6 /M2's, 11 M2/'s, from loc. V87038. Two P4/'s, 1 /P4, 1 /M1, 9 M1/'s, 4 /M2's, 4 M2/'s from loc. V87051. Three P4/'s, 5 /P4's, 5 /M1's, 4 M1/'s, 10 /M2's, 6 M2/'s from loc. V87071. Eight P4/'s, 10 /P4's, 19 /M1's, 18 M1/'s, 14 /M2's, 22 M2/'s from loc. V87072. Six P4/'s, 3 /P4's, 7 /M1's, 4 M1/'s, 9 /M2's, 14 M2/'s from loc. V87074. One P4/, 4 /P4's, 1 /M1, 5 M1/'s, 2 M2/'s from loc. V87077. Three P4/'s, 2 /P4's, 12 /M1's, 6 M1/'s, 8 /M2's, 6 M2/'s from loc. V87098. Two P4/'s, 8 /P4's, 14 /M1's, 10 M1/'s, 9 /M2's, 13 M2/'s from loc. V87151. Twenty five other McGuire Creek localities yielded 1-6 isolated teeth per site.
Localities . V86031, V87028, V87030, V87034, V87035, V87036, V87037, V87038, V87040, V87049, V87051, V87052, V87070, V87071, V87072, V87073, V87074, V87077, V87078, V87084, V87086, V87098, V87101, V87108, V87109, V87114, V87115, V87124, V87151, V87152, V87153, V88038, V88039, and V88041.
Distribution . Upper Hell Creek and lower Tullock formations, Montana, and Polecat Bench Formation (Fort Union Formation), Wyoming (all Puercan).
Discussion and Description . When first proposed, Stygimys included three species formerly referred to Eucosmodon (E. gratus, E. jepseni, E. teilhardi ) and a new species, Stygimys kuszmauli , which was designated as the genotype (Sloan and Van Valen, 1965). Stygimys kuszmauli and S. gratus were reported from the Bug Creek sequence of channel deposits (Bug Creek Anthills, Bug Creek West, Harbicht Hill) in the upper Hell Creek Formation of Montana (Sloan and Van Valen, 1965). Sloan and Van Valen (1965) noted the presence of S. gratus at Harbicht Hill, which was previously known only from Mantua Lentil, Polecat Bench Formation, Wyoming (Jepsen, 1930). The new, smaller species of the genus, S. kuszmauli , was listed as occurring at Bug Creek Anthills (type locality) and Bug Creek West (Sloan and Van Valen, 1965).
The original diagnosis of S. kuszmauli is short and only refers to size differences between it and S. gratus : "This is the smallest species of the genus. Length /P4, 4.6 +/- .3mm (standard deviation of sample); 11 serrations. The specimens of Stygimys from Harbicht Hill are larger than those from Bug Creek Anthills and can be referred to Stygimys gratus " (Sloan and Van Valen, 1965, p. 224). The /P4's of S. gratus from Mantua Lentil have 11 serrations and lengths of 4.8mm and 4.9mm (Jepsen, 1940). These lengths fall within one standard deviation of the mean length of /P4's of S. kuszmauli from Bug Creek Anthills (Table 12). Therefore, the original diagnosis of S. kuszmauli does not distinguish it from S. gratus .
To resolve this problem, the samples of Stygimys in the UCMP collections from Bug Creek Anthills (V65127 and V70201) and Harbicht Hill (V71203) were compared to the small (n=6) sample of Stygimys gratus from Mantua Lentil described by Jepsen (1930; 1940). This analysis indicates that the holotype of S. gratus , PU 13373 a dentary
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
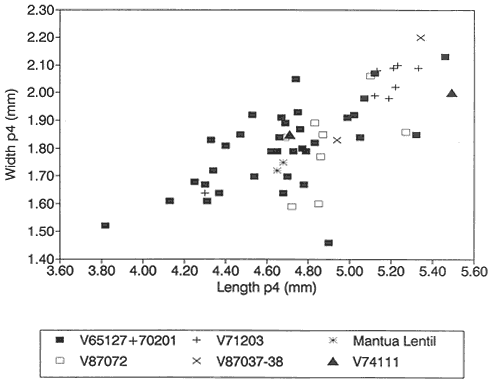
Figure 13
Bivariate plot (length vs. width) of /P4's of Stygimys from Bug Creek Anthills (V65127 and
V70201), Harbicht Hill (V71203), Tedrow Quarry D (V87072), and Up-Up-the-Creek 2 and 3
(V87037-38) (all from the Hell Creek Formation, McCone County, Montana), Worm Coulee
1 (V74111; from the Tullock Formation, Garfield County, Montana), and Mantua Lentil (from
the Polecat Bench Formation, Wyoming). One of the two /P4's from Mantua Lentil
(PU 14496) is encased in sediment, and only length could be measured; its width was
estimated at 1.75 mm, based on the similar length of PU 14419, which has a width of 1.72 mm.
fragment with /M2, represents a different taxon than the remainder of its hypodigm. The holotype (PU 13373) is much smaller than the two other dentary fragments from Mantua Lentil referred to S. gratus . Also, although the /M2 from the holotype is heavily worn, its morphology differs significantly from /M2's of S. kuszmauli from Bug Creek Anthills. Comparison of the holotype of S. gratus with that of Cimexomys hausoi from the Tullock formation of Garfield County, Montana, indicates that these two specimens represent the same taxon. Therefore, PU 13373 becomes the namebearer for that species, now recognized as Cimexomys gratus (further discussion in section on C. gratus ).
Because the holotype of "Stygimys gratus " becomes the namebearer of Cimexomys gratus , the remainder of the Mantua Lentil sample of Stygimys is without species referral. Since the original diagnosis of Stygimys kuszmauli did not distinguish it from the Mantua Lentil sample of Stygimys , comparison of samples from the two type localities, Mantua
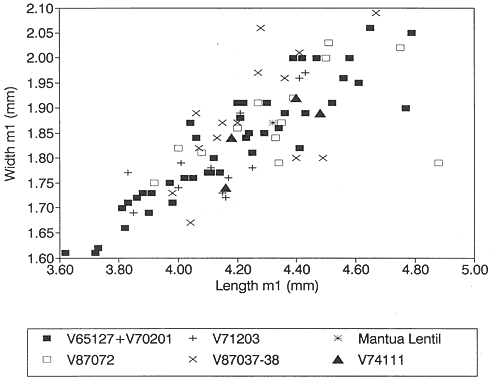
Figure 14
Bivariate plot (length vs. width) of /M1's of Stygimys from various localities (see Figure 13).
Lentil and Bug Creek Anthills, was required. The UCMP sample of Stygimys from Harbicht Hill was included in the analysis because when Stygimys was described initially, the Harbicht Hill sample was referred to "Stygimys gratus " (Sloan and Van Valen, 1965).
/P4: Both /P4's (PU 14496, PU 14419) from Mantua Lentil have 11 serrations. All /P4's from Harbicht Hill have 11 serrations. Three of 36 /P4's from Bug Creek Anthills have 10 serrations, the rest have 11. Based on size, /P4's from Mantua Lentil and Bug Creek Anthills are, on average, nearly identical (Table 12). A bivariate graph of length vs. width indicates that the two /P4's from Mantua Lentil fall within a cluster of specimens of S. kuszmauli from Bug Creek Anthills (Figure 13). The /P4 samples from Harbicht Hill and Bug Creek Anthills have similar ranges in size, but the mean /P4 length of specimens from Harbicht Hill is much larger than that from Bug Creek Anthills (Table 12). However, the difference in mean lengths of /P4's from Harbicht Hill (V71203) and Bug Creek Anthills (V65127 only) was tested statistically, and was not significant (T test: T=-1.23; df 16+ 2.12).
/M1: The single /M1 (PU 14417) from Mantua Lentil has a cusp formula of 6:5. Fifteen percent (7 of 45) of /M1's from Bug Creek Anthills have 6 cusps on the external row, the remainder have seven. All /M1's from Bug Creek Anthills have 5 cusps on the
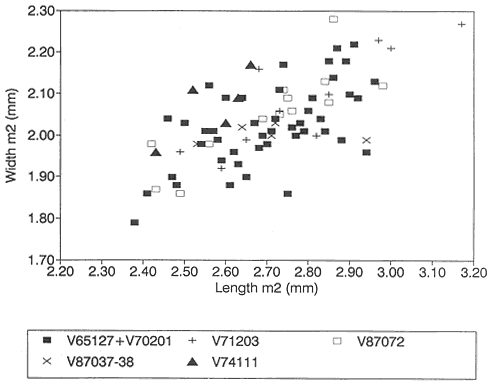
Figure 15
Bivariate plot (length vs. width) of /M2's of Stygimys from various localities (see Figure 13).
internal row. All /M1's from Harbicht Hill (n=10) have 5 cusps on the internal row, and all but one have 7 cusps on the external row; the one exception has 6. Based on size, all three samples are indistinguishable (Table 12). Plotted graphically by length vs. width, /M1's from Harbicht Hill and Mantua Lentil compare closely with those from Bug Creek Anthills (Figure 14).
/M2: The Mantua Lentil sample yields an /M2 (PU 13373) but it is referable to Cimexomys (this report), not Stygimys (contra Jepsen, 1930; sensu Sloan and Van Valen, 1965). Harbicht Hill /M2's (n=10) have a cusp formula of 4:2, as do Bug Creek Anthills /M2's (n=46) except for three, which have 5 cusps on their external rows. Based on size, /M2's from Harbicht Hill are, on average, slightly larger than those from Bug Creek Anthills (Table 12). However, when the two samples are plotted graphically, the small differences in mean length and width are apparently insignificant (Figure 15).
P4/: The Mantua Lentil sample does not contain P4/'s. Harbicht Hill P4/'s (n=3) have a cusp formula of 2-3:7-8:1, and those from Bug Creek Anthills (n=38) 2-3:7-10:1. Based on size, the samples are nearly identical (Table 12). A bivariate length vs. width plot indicates that the Harbicht Hill sample falls completely within the sample from Bug Creek Anthills (Figure 16).
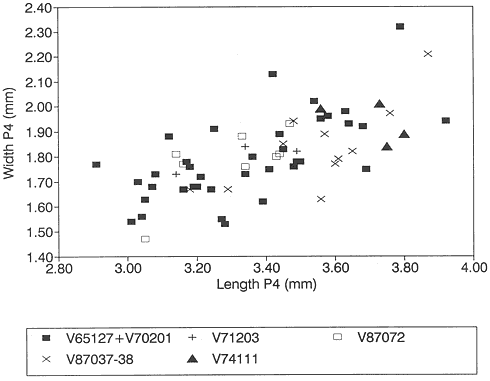
Figure 16
Bivariate plot (length vs. width) of P4/'s of Stygimys from various localities (see Figure 13).
M1/: The two M1/'s (PU 14420A, PU 14420B) from Mantua Lentil are significantly larger than the mean of those from Bug Creek Anthills and Harbicht Hill (Table 12) (PU 14420 refers to both M1/'s, here separated for discussion by A and B designations). However, the total sample from both Harbicht Hill and Mantua Lentil consists of only four specimens (Table 12). From their size alone, these small samples could be interpreted to represent separate species (Figure 17). When larger samples, such as that from Bug Creek Anthills, are included in the analysis, specimens from Mantua Lentil and Harbicht Hill fall within the range of size variation of the Bug Creek Anthills sample. The largest specimens from Bug Creek Anthills are of similar size or larger than those from Mantua Lentil (Figures 17, 18). Therefore, Stygimys kuszmauli and "S. gratus " are not distinguishable by size. In fact, because of the small samples from Harbicht Hill and Mantua Lentil, the strongest argument for recognizing two species of Stygimys on the basis of size would be those from Harbicht Hill and Mantua Lentil, not Bug Creek Anthills and Mantua Lentil.
The cusp formula of M1/'s of Stygimys is highly variable. The two M1/'s from Mantua Lentil have cusp formulas of 7:7:5 (PU 14420A, Figure 18) and 7:7:3 (PU 14420B, not figured), each with 1 or 2 cuspules on the anterior part of the internal cusp row. The two M1/'s from Harbicht Hill have cusp formulas of 8-7:8-7:5-6. The large
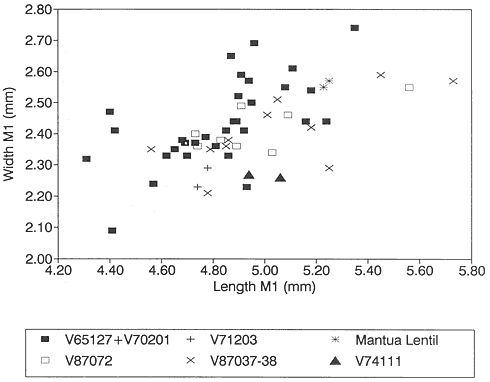
Figure 17
Bivariate plot (length vs. width) of M1/'s of Stygimys from various localities (see Figure 13).
sample of M1/'s from Bug Creek Anthills exhibits cusp formulas of 8-7:8-7:3-8, with the number on the internal row being highly variable. Those from Bug Creek Anthills usually have 3-5 cusps on the internal row, but many cusp counts depend in part on whether tiny cuspules are counted as cusps. The cusp morphology and formula of M1/'s of S. kuszmauli compare closely to those from Mantua Lentil. For example, UCMP 73131 has a cusp formula of 7:7:2 with 4 cuspules (Figure 18), and UCMP 103864 8:8:7 with 1 cuspule (not figured). UCMP 37131 and 103864 are two of the largest M1/'s of Stygimys from BCA and are similar in size to those from Mantua Lentil (PU 14420A and 14420B L: 5.23-5.25, W: 2.55-2.57; UCMP 103864 L: 5.35, W: 2.74; UCMP 73131 L: 5.16, W: 2.44). The samples from Bug Creek Anthills and Harbicht Hill are also similar in morphology. Therefore, on the basis of both size and morphology, samples of M1/'s of Stygimys from Bug Creek Anthills, Harbicht Hill, and Mantua Lentil are not distinguishable.
M2/: The Mantua Lentil sample does not contain M2/'s. Harbicht Hill M2/'s have a cusp formula of 1-2:3:3-4, and those from Bug Creek Anthills 1-3:3:3-4. A third external cusp is present only on a single M2/ from Bug Creek Anthills. Size measurements and a bivariate length vs. width plot show that the samples are very similar (Table 12, Figure 19).
From this analysis, the samples of Stygimys from Bug Creek Anthills, Mantua Lentil, and Harbicht Hill cannot be separated with confidence on the basis of size or
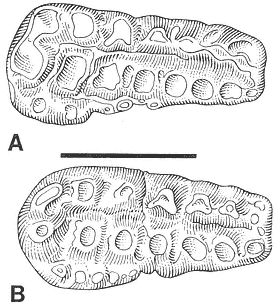
Figure 18
Stygimys kuszmauli Sloan and Van Valen. (A) Isolated right
M1/, UCMP 73131, Bug Creek Anthills (locality V65127),
occlusal view. (B) Isolated right M1/, PU 14420A, Mantua
Lentil, occlusal view. Scale bar = 4 mm.
morphology. Smaller samples of Stygimys (i.e., Mantua Lentil and Harbicht Hill) may exhibit morphological differences, but when larger samples are analyzed (i.e. Bug Creek Anthills), differences appear to have little significance. The morphology of Stygimys is highly variable, which can only be fully appreciated with large samples. The Bug Creek Anthills, Mantua Lentil, and Harbicht Hill samples apparently represent a single, highly variable species of Stygimys that rightfully retains the name S. kuszmauli , the genotypic species.
The sample of Stygimys from the upper Hell Creek Formation at McGuire Creek was compared to those in the UCMP collections from Harbicht Hill (V71203) and Bug Creek Anthills (V65127 and V70201), upper Hell Creek Formation, and Worm Coulee 1 (V74111), lower Tullock Formation. Archibald (1982) studied the Worm Coulee 1 sample and referred it to Stygimys aff. S. kuszmauli . Two of the largest samples from McGuire Creek (V87072, V87037-38) are shown in comparison with Bug Creek Anthills, Harbicht Hill, and Worm Coulee 1 (remeasured for this study) (Figures 13-17, 19). The Stygimys samples from all McGuire Creek sites, Harbicht Hill, Bug Creek Anthills, and Worm Coulee 1 exhibit a wide range of morphological variation, but these differences fade when large samples are analyzed. Fossils from these samples are nearly identical in size and morphology (Tables 12, 13, Figures 13-17, 19), and probably belong to a single species. Using these comparisons and the absence of distinguishing characters, I refer the samples from McGuire Creek sites and Worm Coulee 1 to Stygimys kuszmauli .
Fox (1989) described a new species of Stygimys, S. cupressus from the Long Fall
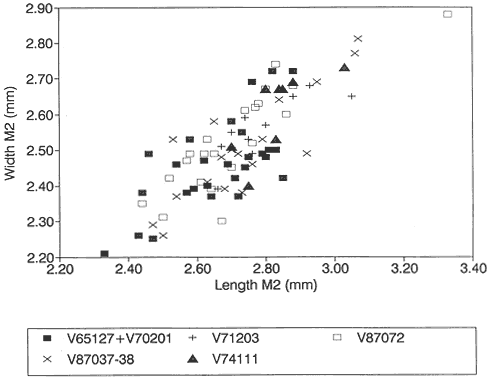
Figure 19
Bivariate plot (length vs. width) of M2/'s of Stygimys from various localities (see Figure 13).
locality of the Ravenscrag Formation in Saskatchewan. The species is diagnosed by the /P4, whose cutting edge has a symmetrical arc with a long, low, inclined anterior edge leading to the first serration (Fox, 1989). Also, specimens in the University of Alberta collections reveal that the /P4 of S. cupressus (n=4) is longer than that of S. kuszmauli (n=9) (Fox, 1989). However, lengths of /P4's from Long Fall (4.9mm-5.4mm) fall within the upper part of the range of variation of the large UCMP sample from Bug Creek Anthills (n=34; see Figure 13 and Table 12). A direct comparison of a large sample of S. kuszmauli from Bug Creek Anthills and the Long Fall sample of S. cupressus would provide an interesting test as to the validity of the latter species because of the highly variable morphology of S. kuszmauli .
The morphology of Stygimys kuszmauli has been described in detail (Archibald, 1982, formerly Stygimys aff. S. kuszmauli ), but some new information is provided by the fossils from McGuire Creek and Harbicht Hill.
Almost all /P4's from McGuire Creek (n=40) and Harbicht Hill (n=8) have 11 serrations. Three of these from the large McGuire Creek sample have only 10 serrations (UCMP 134739, loc. V87072; UCMP 132666, loc. V87037; UCMP 132803, loc. V87078). The P4/'s from Harbicht Hill and McGuire Creek have a cusp formula of 2-3:6-10:1.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
McGuire Creek and Harbicht Hill /M2's have a cusp formula of 3-5:2. Ninety percent of the /M2's (n=60) have 4 external cusps. One specimen (UCMP 134725, loc. V87072) has a large fifth external cusp, which is weakly separated from the fourth external cusp by a shallow lingual groove. The M2/'s have a cusp formula of 1-3:3:3-5. Specimens with 4 or 5 internal cusps are rare (5 of 60). When an additional cusp is developed, it is either anterior to and not well separated from the first internal cusp or it is developed posterior to the third internal cusp and deflected medially to occupy a position at the posterior end of the medial valley. One specimen (UCMP 132138, loc. V87151) has cusps developed in both of these positions, giving the tooth 5 cusps in the internal row.
The /M1's have a cusp formula of 6-8:5. When an eighth external cusp is present, it is developed as a robust cusp on a labial bulge within the notch separating cusps 4 and 5, or as a small cusp on a low ridge that extends posteriorly from cusp 4.
The M1/'s of Stygimys kuszmauli from McGuire Creek and Harbicht Hill have a cusp formula of 7-8:7-8:3-7. When cusp 8 is present on the external or medial row, it is small and positioned on the anterior margin of the tooth. Three to 7 cusps comprise the internal row. When more than 5 cusps are present, the internal row extends anteriorly to, or slightly past, the midline of the tooth. Cusps on the internal row become progressively smaller anteriorly, with cusps 6 and 7 more appropriately described as tiny cuspules. This is similar to the condition found in M1/'s of S. kuszmauli from Mantua Lentil (formerly S. gratus ).
Family TAENIOLABIDIDAE Granger and Simpson, 1929
Catopsalis Cope, 1882a
Comments: Catopsalis is a paraphyletic taxon (Simmons and Desui, 1986) known from Upper Cretaceous strata in Asia (Kielan-Jaworowska and Sloan, 1979) and Puercan to Tiffanian strata in North America (Archibald et al., 1987; Archibald and Lofgren, 1990). Two species of the genus, C. joyneri and C. alexanderi , have been reported from eastern Montana (Sloan and Van Valen, 1965; Middleton, 1982). C. joyneri is best known from the Bug Creek sequence of channel fills (Bug Creek Anthills, Bug Creek West, Harbicht Hill) in the upper Hell Creek Formation of McCone County, Montana. The type of C. alexanderi comes from the Alexander locality in the Denver Formation, Arapahoe County, Colorado. The hypodigm of C. alexanderi included specimens originally referred to C. foliatus (see Middleton, 1982, p. 1198). Two specimens also referred to C. alexanderi in the hypodigm were collected from the Tullock Formation of Garfield County, Montana. These fossils (UCMP 116954, M1/ fragment, UCMP loc. V74111; UCMP 124404, M2/, UCMP loc. V74110) were originally referred to Catopsalis cf. C. foliatus (see Archibald, 1982).
Species of Catopsalis display certain evolutionary trends when set in decreasing chronologic order (Kielan-Jaworowska and Sloan, 1979; Middleton, 1982). One of these is a gradual increase in size: Catopsalis joyneri is smaller than C. alexanderi , which in turn is smaller than C. foliatus .
Catopsalis joyneri , and the distinctly larger C. alexanderi , are both present at McGuire Creek, but never at the same locality. This might be a factor of the small sample size (n=10), or may reflect temporal differences between sites.
Catopsalis joyneri Sloan and Van Valen, 1965
Table 14
Catopsalis joyneri Sloan and Van Valen, 1965, p. 225.
Type . UMVP 1494, right maxilla with complete palate, M1/, roots of P4/, alveoli of P3/ and M2/.
Type locality . Bug Creek Anthills, Hell Creek Formation.
Referred specimens . One P4/ from loc. V87071. One M2/, 1 /M1 from loc. V87151. One M2/ from loc. V87153.
Localities . UCMP locs. V87071, V87151, and V87153.
Distribution . Upper Hell Creek Formation, Montana (Puercan); possibly upper Frenchman and Ravenscrag formations, Saskatchewan (both Puercan).
Description . In 1965, Sloan and Van Valen published a brief description of C. joyneri based on fossils collected at Bug Creek Anthills, McCone County, Montana. A more complete description of the species has yet to appear. Therefore, a brief description of the small McGuire Creek sample (n=4) is given below.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Dental measurements of C. joyneri from McGuire Creek are presented in Table 14. The UCMP sample of C. joyneri from Bug Creek Anthills was measured, and these data are also given in Table 14 for comparative purposes.
The single P4/ (UCMP 133167) referable to C. joyneri is heavily worn on its medial crest, making determination of the cusp formula impossible (1:5?:?). The single external cusp is small and positioned on a well developed cingulum on the posterolabial edge of the tooth. The medial row probably had 5 cusps, but only cusps 1-3 remain. The medial row appears to be similar to that of the larger species of Catopsalis, C. alexanderi (see Middleton, 1982, p. 1201). The posterolingual part of the tooth is heavily worn and any trace of internal cusps has been removed. A short but distinct cingulum is present labial to medial cusps 4-5?.
A moderately worn /M1 (UCMP 132181) has a cusp formula of 5:4 and is similar in size and morphology to /M1's in the UCMP sample from Bug Creek Anthills. UCMP 133567, a slightly worn M2/, has a cusp formula of 1:3:3. Its internal cusps are subequal in size. In the medial row, the anterior cusp is small and transversely elongated, and the posterior cusp is the largest. The single external cusp, on the anterolabial edge of the tooth, is narrow and ridge-like, with its long axis oriented anterolingual-posterolabial. This cusp is positioned on the anterolabial edge of the tooth. The other M2/, UCMP 132180, is heavily worn and offers no additional morphological information.
Catopsalis alexanderi Middleton, 1982
Table 15
Catopsalis alexanderi Middleton, 1982, p. 1198.
Type . UCM 34979, right lower jaw with I, /P4, /M1-2.
Type locality . Alexander locality (UCM 77267), Denver Formation, Colorado.
Referred specimens . One M2/, UCMP 136095, from loc. V71203. One /M2, UCMP 132478, from loc. V87034. One M1/, UCMP 132659, from loc. V87037.
Localities . UCMP locs. V71203, V87034, and V87037.
Distribution . Denver Formation, Colorado; Polecat Bench Formation, Wyoming; upper Hell Creek and lower Tullock formations, Montana (all Puercan).
Description . The three specimens referred to C. alexanderi are similar in size to those in Middleton's (1982, tables 1, 2) sample and are significantly larger than those of C. joyneri
| ||||||||||||||||||||||||||||||
from Bug Creek Anthills (Tables 14, 15). Morphologically, these specimens are similar to those described by Middleton (1982), and only a few comments are necessary.
UCMP 132659, an unworn M1/, is missing the anterior portion of the external cusp row because of breakage, and only 5 cusps are preserved. However, a proportional comparison to complete M1/'s of C. joyneri reveal that UCMP 132659 probably had at least 7 cusps. The middle and internal rows have 8 cusps each. The internal row extends anteriorly about 90% of the total length of the tooth. UCMP 132478, a moderately worn /M2, is missing the extreme posterolabial corner of the tooth. From comparisons to complete /M2's of C. joyneri from Bug Creek Anthills, it appears that the length of UCMP 132478 (Table 15) is close to the original maximum length. This specimen has a cusp formula of 3:2+. The internal row has a small narrow anterio-posteriorly orientated cuspule on the postero-labial margin of the tooth.
The small UCMP collection from Harbicht Hill (V71203) contains one isolated tooth, an M2/ (UCMP 136095), referable to Catopsalis (Table 15); it is much larger than that of C. joyneri (Table 14). Based on size and morphology (cusp formula 1:3:3), UCMP 136095 is referable to C. alexanderi .
The presence of C. alexanderi at Harbicht Hill and McGuire Creek are new records of the species in the upper Hell Creek Formation. Previously, only C. joyneri was listed as present at Harbicht Hill (Sloan and Van Valen, 1965). Because C. alexanderi was described more recently (Middleton, 1982) than Sloan and Van Valen's (1965) Harbicht Hill faunal list, it is possible that either both species or only C. alexanderi is present at Harbicht Hill.
Catopsalis sp. indet.
Table 16
Referred specimens . One /I from loc. V87033. One /I from loc. V87035. One /I fragment from loc. V87070. One /I fragment from loc. V87071. One /Mx fragment from loc. V87074.
Localities . UCMP locs. V87033, V87035, V87070, V87071, and V87074.
Description . These four lower incisors and a partial molar are referable to Catopsalis with reasonable confidence, but are too fragmentary for identification to species level.
Lower incisors of Catopsalis alexanderi and C. joyneri are distinguished by size (Middleton, 1982). Lower incisors of Catopsalis from McGuire Creek appear to be referable to C. alexanderi as shown by comparison of measurements from Table 16 and Table 2
| ||||||||||||||||||||||||
of Middleton (1982, p. 1201). However, the size range and mean dimensions of incisors referred to C. joyneri were not presented in the original diagnosis of the species (Sloan and Van Valen, 1965). Therefore, the incisors listed in Table 16 are not referred to C. alexanderi , although this possibility appears most likely, based on size.
Suborder, incertae sedis
Family CIMOLOMYIDAE (Marsh, 1889b)
Sloan and Van Valen, 1965
Meniscoessus Cope, 1882b
Meniscoessus robustus (Marsh, 1889a) Osborn, 1891
Table 17
Dipriodon robustus Marsh, 1889a, p. 85 (see Archibald, 1982, p. 75, for synonymies).
Type . YPM 11234, right /M2 (Marsh 1889a, pl. II, figs. 13-15).
Type locality . Mammal locality no. 2 of Lull (1915), UCMP loc. V5815, Lance Formation, Wyoming.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Referred specimens . Four M1/'s, 4 M2/'s, 1 /P4, 3 /M1's, 1 /M2 from loc. V85092. Three I's, 3 P4/'s, 1 M1/, 7 M2/'s, 2 /P4 fragments, 7 /M1's, 5 /M2's, and 2 Mx fragments from other UCMP localities.
Localities . UCMP locs. V84151, V85092, V86031, V87030, V87034, V87035, V87037, V87038, V87071, V87072, V87074, V87084, V87098, V87151, and V88037.
Revised diagnosis . Archibald, 1982, p. 75.
Distribution . St. Mary River Formation, Alberta (Edmontonian); Hell Creek Formation, South Dakota, and Lance Formation, Wyoming (both Lancian); Frenchman Formation, Saskatchewan (Lancian, in part); Hell Creek Formation, Montana (Lancian and *Puercan); possibly upper Frenchman Formation and Ravenscrag Formation, Saskatchewan (both *Puercan).
Description . The morphology of Meniscoessus robustus has been adequately described by Clemens (1964) and Archibald (1982). McGuire Creek specimens referred to M. robustus are similar in size and morphology to fossil material described in those works, and no additions are necessary. Dental measurements are given in Table 17.
Comments . Fossils referred to Meniscoessus robustus come from both Lancian and Puercan localities in the upper Hell Creek Formation at McGuire Creek. All the Lancian specimens are from UCMP locality V85092. The remaining teeth were recovered from 14 widely scattered Puercan localities. The presence of Meniscoessus at these Puercan sites probably is the result of reworking. Alternatively, the presence of two morphologically distinct samples, one each from Puercan and Lancian sites, would suggest that Meniscoessus might have survived into the Puercan. To test this hypothesis, the entire McGuire Creek sample of Meniscoessus was measured and then divided into the two bulk age groupings (Lancian and Puercan). Comparison of these admittedly small samples gave no indication of significant size or morphological differences (Table 17). The Puercan specimens are, on average, slightly larger, but almost all of these fossils fall within the range of variation displayed by M. robustus from the Lance Formation of Wyoming (Clemens, 1964, table 10) and the Lancian part of the Hell Creek Formation of Montana (Archibald, 1982, table 13). The size of dental elements of M. robustus appears to be highly variable, and until collection of a larger sample from McGuire Creek shows evidence of a distinct bimodality between Lancian and Puercan specimens of Meniscoessus , it appears only that one species is present.
Family ?CIMOLOMYIDAE
Essonodon Simpson, 1927a
Table 18; Figure 20
Essonodon browni Simpson, 1927a, p. 2.
Type . AMNH 14410, an isolated right /M2 (Simpson, 1927a, fig. 1).
Type locality . Near the head of the East Fork of Crooked Creek, Hell Creek Formation, Montana.
Referred specimen . One /M2, UCMP 132182, from loc. V87151.
Locality . UCMP loc. V87151.
Distribution . Kirkland Formation, New Mexico (Lancian?); Hell Creek Formation,
| |||||||||||||||||||||||||||||||||||
Montana (Lancian and *Puercan); possibly Frenchman Formation, Saskatchewan (Lancian, in part).
Revised diagnosis . Archibald, 1982, p. 89-90.
Description . The single tooth referred to Essonodon browni from McGuire Creek is a moderately worn /M2 (UCMP 132182) with a cusp formula of 3:2. This specimen is similar in size and morphology to the holotype (AMNH 14410, an isolated /M2) and to the few other /M2's known of the species (Table 18). UCMP 132182 (Figure 20) differs slightly from the holotype in development and orientation of the posterior external cusp, which is smaller, located more posteriorly, and situated on the extreme posterolabial edge of the tooth. A well-developed, transversely oriented ridge, which is enhanced by the moderate state of wear of the tooth, forms the posterior margin of the tooth and connects the external and internal cusps. On the holotype, this ridge is less marked and the posterior external cusp is larger and situated more anteriorly (Simpson, 1927a).
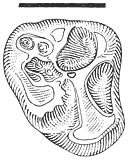
Figure 20
Essonodon browni Simpson.
Isolated left /M2, UCMP
132182, locality V87151,
occlusal view. Scale
bar = 2 mm.
Suborder and Family, incertae sedis
Cimexomys Sloan and Van Valen, 1965
Cimexomys minor Sloan and Van Valen, 1965
Tables 19-20
Cimexomys minor Sloan and Van Valen, 1965, p. 221.
Type . SPSM 62-2115, left mandible with /P3-4, and alveoli for /I, /M1, and /M2 (Sloan and Van Valen, 1965, fig. 2).
Type locality . Bug Creek Anthills, Hell Creek Formation, Montana.
Referred specimens . One /P4 from loc. V87030. Three M1/'s, 3 /P4's, from loc. V87037. Eight M1/'s, 1 /P4, 2 P4/'s from loc. V87038. One M1/ from loc. V87051. One M1/ fragment from loc. V87071. Seven M1/'s, 5 /P4's, 2 P4/'s from loc. V87072. Four M1/'s, 2 /P4's from loc. V87074. One M1/ from loc. V87077. One P4/ from loc. V87091. One M1/, 2 /P4's from loc. V87098.
Localities . UCMP locs. V87030, V87037, V87038, V87051, V87071, V87072, V87074, V87077, V87091, and V87098.
Distribution . Lance Formation, Wyoming (Lancian); Hell Creek Formation, Montana (Lancian and Puercan); lower Tullock Formation, Montana, and Ravenscrag Formation, Saskatchewan (both Puercan).
Description . The McGuire Creek sample of Cimexomys minor is limited to /P4's, P4/'s, and M1/'s. The abundant presence of C. minor at some sites (especially locs. V87072, V87037, and V87038) would indicate that /M1's, /M2's, and M2/'s are undoubtedly also present in the collections from these and other localities at McGuire Creek. Unfortunately, current knowledge does not allow teeth of C. minor to be distinguished from smaller species of Mesodma , unless found in association with /P4's, P4/'s, or M1/'s (Archibald, 1982). Therefore, the large Mesodma sample from McGuire Creek (n>2200) must include a total number of /M1's, /M2's, and M2/'s of Cimexomys minor roughly equal to the number of identifiable /P4's, P4/'s, and M1/'s (n=45).
Specimens referred to Cimexomys minor are similar to those described by Sloan and Van Valen (1965), Clemens (1964, 1973), and Archibald (1982), and only a few comments are necessary. M1/'s from McGuire Creek have a cusp formula of 4-5:6-7:1. Twenty percent (n=5) of them have 7 cusps in the medial row, and four of these teeth also have 5 cusps in the external row. The internal cusp is elongated antero-posteriorly and forms a short ridge, which does not extend to the midline of the tooth. In a few unworn specimens, this internal ridge can be subdivided into 2 or 3 weakly developed cuspules. /P4's have 8 serrations with 5-6 internal and external ridges, which are not developed on the first or last serrations. All P4/'s from McGuire Creek have a cusp formula of 3:5:2.
Dental measurements of C. minor from the localities which form the majority of the McGuire Creek sample are presented in Table 19. Fossils from McGuire Creek are similar in size to those from the Lance Formation of Wyoming (UCMP loc. V5711) (Clemens, 1973, table 8) and the lower Tullock Formation of Montana (UCMP loc. V74111) (Archibald, 1982, table 15). M1/'s in the UCMP collections from the type locality of the species (Bug Creek Anthills, UCMP loc. V65127) also compare closely in size to those from McGuire Creek (Table 20).
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||
Cimexomys gratus Jepsen, 1930
Tables 21-22; Figures 21-22
Cimexomys hausoi Archibald, 1982, p. 105.
Type . PU 13373, incomplete left ramus with /M2, and alveoli of I, /P4, and /M1 (Jepsen, 1930, pl. IV, fig. 8).
Type locality . Mantua Lentil, Polecat Bench Formation (Fort Union Formation), Wyoming.
Referred specimens . One /P4, 4 P4/'s, 3 /M1's, 2 M1/'s, 2 /M2's, 1 M2/, from loc. V87037. One /P4, 1 /M1, 2 M1/'s, 5 M2/'s, from loc. V87035. Three P4/'s, 2 /P4's, 3 /M1's, 6 M1/'s, 3 /M2's, from loc. V87038. Two P4/'s, 4 /M1's, 1 M1/, 2 /M2's, 6 M2/'s, from loc. V87151. One P4/, 1 /M1, 1 M1/, 3 M2/'s, from loc. V87051. One P4/, 1 /P4, 1 /M1, 6 M1/'s, 2 M2/'s, from loc. V87098. One /P4, 2 /M1's, 2 M1/'s, 3 /M2's, 4 M2/'s, from loc. V87071. One /P4, 4 P4/'s, 1 /M1, 7 M1/'s, 7 M2/'s, 3 /M2's, from loc. V87074. Four /P4's, 8 P4/'s, 7 /M1's, 10 M1/'s, 5 /M2's, 8 M2/'s, from loc. V87072. One /M1, 1 M1/, from loc. V65127. Two M1/'s from loc. V71203. Seven isolated premolars and molars from other UCMP localities.
Localities . UCMP locs. V65127, V71203, V87028, V87030, V87035, V87037, V87038, V87051, V87052, V87071, V87072, V87074, V87077, V87098, V87101, V87114, and V87151.
Distribution . Upper Hell Creek and lower Tullock formations, Montana (both Puercan); Polecat Bench Formation (Fort Union Formation), Wyoming, and possibly Ravenscrag Formation, Saskatchewan (both Puercan).
Discussion . During the course of study of the Stygimys gratus sample from Mantua Lentil, which was originally described by Jepsen (1930, 1940), it was apparent that the holotype and the remainder of the hypodigm represented two different taxa. The hypodigm, excluding the holotype, is indistinguishable from Stygimys kuszmauli from Bug Creek Anthills and is referred to that species (see discussion of S. kuszmauli ). The holotype of S. gratus , a fragmentary dentary with a heavily worn /M2 (Figure 21), is similar to the holotype of Cimexomys hausoi from Worm Coulee 1 (V74111), lower Tullock Formation, Montana (Archibald, 1982, figure 36), where comparisons can be made. Measurement of the depth of the dentary below the /M2 (measured from alveolar rim of the /M2 to ventral edge of the dentary) indicates that the holotype of Stygimys gratus (PU 13373: 4.2mm) is similar to the holotype of Cimexomys hausoi (UCMP 117000: 3.9mm). These two specimens are much smaller than the two other dentary fragments from Mantua Lentil referred to Stygimys gratus (PU 14418: 7.7mm; PU 14419: 6.5mm). Also, the /M2's of these holotypes are similar in size (Cimexomys hausoi UCMP 117000: L: 2.32, W: 1.86; Archibald, 1982, table 16; Stygimys gratus PU 13373: L: 2.37, W: 1.85) and have cusp formulas of 4:2. However, based on size, PU 13373 falls within the range of variation of /M2's of S. kuszmauli from Bug Creek Anthills (L: 2.38-2.96; W: 1.79-2.22). Therefore, comparison of PU 13373 to worn /M2's of Cimexomys hausoi and Stygimys kuszmauli from their respective type localities is needed to eliminate the possibility that PU 13373 represents a small individual of the latter species.
The /M2 of PU 13373 is similar to worn /M2's of Cimexomys hausoi from Worm Coulee 1 (V74111) in the following ways: (1) /M2's of Stygimys kuszmauli initially exhibit the heaviest wear on the internal cusp row, while those of Cimexomys hausoi
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
exhibit heaviest wear on the external cusp row first (Archibald, 1982). (2) Transverse grooves between cusps of Stygimys kuszmauli are wide and broad, especially those between the internal cusps and the two anterior external cusps. /M2's of Cimexomys hausoi have narrow deep grooves between cusps on both cusp rows, which are deepest medially on each row. (3) The internal cusps of Stygimys kuszmauli merge to form a longitudinal ridge only after extreme wear, because a deep, broad groove separates the two cusps. After moderate wear, the transverse grooves between cusps of Cimexomys hausoi are removed from the lingual edge of the internal row and labial edge of the external row, and cusps in each row merge to form two longitudinal ridges. (4) An irregular series of deep pits is present in the enamel on the occlusal surface of /M2's of Cimexomys hausoi . Pitting of the enamel is poorly developed or absent in Stygimys kuszmauli . (5) The anterior end of /M2's of Cimexomys hausoi is flattened and has a straight anterior face to which the posterior end of the /M1 abuts. In contrast, the first internal cusp of Stygimys kuszmauli is swollen and rounded, and the anterior face is not flattened to form a straight anterior surface.
The holotypes of Stygimys gratus and Cimexomys hausoi are virtually identical, and these specimens represent a single species. Neither Stygimys gratus nor Cimexomys hausoi are the genotypic species of their respective genera, and both genera were described
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
in the same work (Sloan and Van Valen, 1965). Thus, the holotype of Stygimys gratus (PU 13373) replaces the holotype of Cimexomys hausoi (UCMP 117000) and becomes the namebearer of that species, now recognized as Cimexomys gratus .
During the measurement of M1/'s of Cimexomys from Bug Creek Anthills, it became evident that there were two species present in the sample. Most specimens are small and can be referred to C. minor (see Table 20), but six teeth are much larger. Based on cusp and overall tooth morphology, these six teeth are indistinguishable from those referred to C. gratus from Worm Coulee 1 (V74111). Based on size, most fall within the range of variation displayed by C. gratus (Table 21, Figure 22). The specimens from Bug Creek Anthills form a cluster near the two smallest specimens from Worm Coulee 1, the type locality of C. gratus (Figure 22). Specimens from Bug Creek Anthills would presumably be referred to C. gratus if they were collected from Worm Coulee 1. As an example, the range in variation of the sample of C. gratus from V87037-38 is nearly as large as both the Bug Creek Anthills and Worm Coulee 1 samples in combination (Figure 22, Table 21). In any case, evidently C. gratus and C. minor are both present at Bug Creek Anthills. In Sloan and Van Valen's (1965) brief description of the fauna at Bug Creek Anthills, no mention is made of a larger species of Cimexomys .
Although improbable, a third species of Cimexomys might be present in the upper Hell Creek Formation of McCone County, Montana. In the original description of C. minor , Sloan and Van Valen (1965) reported a second and larger species of Cimexomys from Harbicht Hill and referred a single specimen (PU 14999) from Mantua Lentil (Polecat Bench Formation, Wyoming) to it. However, they did not name or describe the species. Subsequently, Archibald (1982) described a large species, C. gratus (formerly C. hausoi ), from the lower Tullock Formation of Garfield County, Montana.
Comparison of C. gratus to the unnamed species at Harbicht Hill was not possible, because the original specimens from Harbicht Hill could not be located (Archibald, 1982). Examination of the specimen from Mantua Lentil (PU 14999) by Middleton (1983) and comparison with Archibald's description and measurements of C. gratus revealed no significant differences (Middleton, 1983, p. 150-152). Also, the small UCMP collection from Harbicht Hill (V71203) contains two M1/'s (UCMP 136096 and 136097) referable to Cimexomys . These teeth were compared to M1/'s of C. gratus from Worm Coulee 1 (V74111) and found to be morphologically indistinguishable (see Table 21 and
Figure 21
Cimexomys gratus Jepsen. Left dentary fragment with /M2, and alveoli of /I, /P4, and /M1,
PU 13373 (holotype), Mantua Lentil, occlusal view. Scale bar = 2mm.
Figure 22; both samples have cusp formulas of 5:6:1). Therefore, the unnamed large species of Cimexomys at Harbicht Hill mentioned by Sloan and Van Valen (1965) almost certainly is C. gratus .
Description . Fossils of C. gratus from McGuire Creek are similar to those described by Archibald (1982). P4/'s from McGuire Creek have a cusp formula of 3-4:5-6:2-3, with 3 internal cusps present in only one specimen. /P4's have 8-9 serrations, with the majority (9 of 11) having nine. Specimens with 8 serrations have 6 internal and external ridges, none of which are developed from the first or last serration. Fossils with 9 serrations exhibit more variation in ridge development with 6-8 external and internal ridges present. The ultimate serration never has a ridge. In specimens with 6 external or internal ridges, the penultimate serration lacks a ridge. Three specimens have a ridge count of 8 (external and internal), because the first serration has a very short, weakly developed ridge.
M1/'s have a cusp formula of 4-5:6:1 (R-4). An anteriorly positioned fifth external cusp is well developed when present (65% of 40 teeth), and all specimens have 6 medial cusps. /M1's have a cusp formula of 6-7:4, with 6 external cusps present in only a single specimen. M2/'s have a cusp formula of 1:3:4-5, with a single tooth having a fifth internal cusp. /M2's have a cusp formula of 4:2. Dental measurements are given in Table 22.
Archibald (1982), analyzing fossils from the Worm Coulee 1 (UCMP loc. V74111) locality in the lower Tullock Formation of Garfield County, Montana, noted that most of the differences between C. minor and C. gratus (formerly C. hausoi ) are attributable to size. This is also true for the McGuire Creek sample of the genus, although three morphological differences are worth noting. A few M1/'s of C. minor from McGuire Creek have 7 medial cusps instead of six. M1/'s of C. gratus from McGuire Creek always have 6 medial cusps. Also, in many M1/'s of C. gratus the fifth anterior external cusp is large and well separated from the fourth. When a fifth anterior external cusp is present in C. minor , it is small and not well separated from the fourth external cusp. /P4's of C. minor have 8 serrations while /P4's of C. gratus have 8-9 serrations with most having nine.
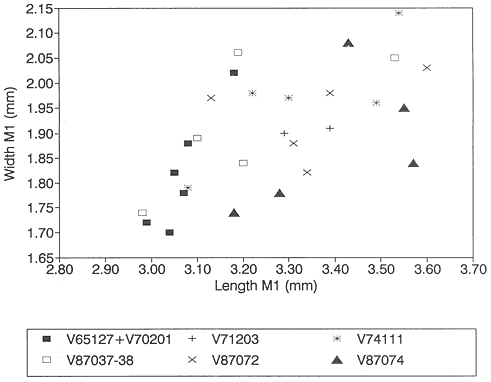
Figure 22
Bivariate plot (length vs. width) of M1/'s of Cimexomys from Bug Creek Anthills (V65127
and V70201), Harbicht Hill (V71203), Up-Up-the-Creek 2 and 3 (V87037-38), Tedrow
Quarry D (V87072) (all from the Hell Creek Formation, McCone County, Montana), and
Worm Coulee 1 (V74111; from the Tullock Formation, Garfield County, Montana).
Subclass THERIA Parker and Haswell, 1897
Infraclass TRIBOSPHENIDA McKenna, 1975
Supercohort MARSUPIALIA (Illiger, 1811) Cuvier 1817
Cohort ALPHADELPHIA Marshall, Case, and Woodburne, 1990
Order PERADECTIA Marshall, Case, and Woodburne, 1990
Superfamily PERADECTOIDEA (Crochet, 1979) Marshall, Case, and Woodburne, 1990
Family PERADECTIDAE (Crochet, 1979) Marshall, Case, and Woodburne, 1990
Subfamily CAROLOAMEGINIIDAE Ameghino, 1901
Glasbius Clemens, 1966
Glasbius twitchelli Archibald, 1982
Tables 23-24
Glasbius twitchelli Archibald, 1982, p. 137
Type . UCMP 115853, an isolated right M3/.
Type locality . UCMP loc. V73087, Hell Creek Formation, Montana.
Referred specimens . Two /Mx fragments, 1 /M2, and 1 Mx/ fragment from loc.
| ||||||||||||||||||||||||||||||||||||||||||||||||
V87035. Two /Mx's, 1 M2/, and 1 Mx/ fragment from loc. V87037. Dentary fragment with /P4, /M3-4, UCMP 133069, and 2 /M2's from loc. V87038. Maxillary fragment with M2-3/ UCMP 1347101, 1 /M3, and 1 M3/ from loc. V87072. One M2/, 1 /Mx, and 1 Mx/ from loc. V87074.
Localities . UCMP locs. V87035, V87037, V87038, V87072, and V87074.
Distribution . Hell Creek Formation, Montana (Lancian and *Puercan); Frenchman Formation, Saskatchewan (Lancian, in part).
Comments . Using fossils from the Lance Formation, Wyoming, and a mandibular fragment from the Hell Creek Formation, Montana, Clemens (1966) described a new monogeneric subfamily of didelphids, the Glasbiinae, composed of one species, Glasbius intricatus . Archibald (1982), when analyzing mammal faunas from the Hell Creek Formation in Montana, noted several minor morphological characters that could be used to distinguish a new species of Glasbius . On the basis of these characters, Archibald (1982) erected Glasbius twitchelli and referred the entire Hell Creek Formation sample to it. As a result, G. intricatus was restricted to the Lance Formation, Wyoming.
The minor morphological features used to distinguish G. twitchelli from G. intricatus are also consistently present on fossils from McGuire Creek and clearly support Archibald's (1982) decision to erect a new species for the Hell Creek sample. Upper and lower molars from McGuire Creek referred to G. twitchelli are larger than G. intricatus (compare Tables 23, 24; with Clemens, 1966, table 6). Also, in all specimens from McGuire Creek, M2/ and M3/'s lack C cusps, M3's only possess a single D cusp, and /M2's and /M3's consistently have one (/M2) or two (/M3) well-developed basal cingulum cusps. These are the main characters used to distinguish G. twitchelli from G. intricatus (Archibald, 1982).
Description . All specimens of Glasbius from McGuire Creek are referred to G. twitchelli . The McGuire Creek sample is similar in morphology and size (Tables 23, 24) to fossils previously described by Archibald (1982) from Montana. The only difference of note is that /M3's from McGuire Creek are slightly wider (Table 24).
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Subfamily ALPHADONTINAE
Marshall, Case, and Woodburne, 1990
Alphadon Simpson, 1927b
Comment . Systematic studies of Alphadon available in 1990 led to the recognition of A. rhaister, A. marshi , and A. wilsoni as present in the upper Hell Creek Formation at McGuire Creek. A fourth, unnamed species of Alphadon , intermediate in size between A. marshi and A. wilsoni , briefly described by Johnston and Fox (1984) and Fox (1989) from the Long Fall (Ravenscrag Formation) and Wounded Knee (Frenchman Formation) local faunas in Canada, was also present at McGuire Creek. Recently, Storer (1991) named a new species of Alphadon, A. jasoni , from the Gryde Local Fauna (Late Cretaceous), Saskatchewan, and referred the entire Alphadon sample from Wounded Knee to it. Also, Storer (1991) referred an upper molar from Bug Creek Anthills to A. jasoni , but did not mention if the Alphadon sample from the Long Fall locality contained A. jasoni .
More recently, Zerina Johanson (1991) presented a preliminary revision of Alphadon marshi and Alphadon wilsoni in which two upper molar morphologies are recognized, based on various characters including shape of the centrocrista and position of stylar cusp C. The first is similar to A. marshi and includes the type of A. wilsoni (A. wilsoni becomes the junior synonym of A. marshi ) and the other is similar to the morphology of A. jasoni described by Storer (1991). Judd Case is also working on a major systematic revision of Alphadon (pers. comm., 1991). Therefore, specimens from McGuire Creek presently referred to A. wilsoni or Alphadon species indeterminate may be eventually referred to A. jasoni or A. marshi . Until final revisions by Johanson and Case are available, specimens from McGuire Creek will be referred to A. wilsoni and
Alphadon species indeterminate, but the species A. "wilsoni" will be recognized in quotations pending its formalized synonomy with A. marshi .
Alphadon "wilsoni" Lillegraven, 1969
Tables 25-26
Alphadon wilsoni Lillegraven, 1969, p. 39.
Type . UA 3681, fragmentary right maxilla with M2-3/ (Lillegraven, 1969, fig. 19-4).
Type locality . Loc. KUA-1, Scollard Formation, Alberta.
Referred specimens . One M1/ from UCMP loc. V87037. Two M1/'s, 1 M2/, 1 /Mx, from UCMP loc. V87072. One /Mx from UCMP loc. V87073. Two /Mx's, 1 /M4 from UCMP loc. V87074. One /M1 from UCMP loc. V87101.
Localities . UCMP locs. V87037, V87072, V87073, V87074, and V87101.
Distribution . Scollard Formation, Alberta, and Lance Formation, Wyoming (both Lancian); Hell Creek Formation, Montana (Lancian and *Puercan); possibly Frenchman Formation, Saskatchewan (Lancian, in part).
Description . All upper molars referred to A. "wilsoni" from McGuire Creek have a large stylar C cusp that is well separated from cusp B. This development of the stylar cusps distinguishes A. "wilsoni" from Protalphadon lulli , which has a small stylar cusp C that is not well separated from cusp B (Clemens, 1966; Cifelli, 1990). In size and proportions, the McGuire Creek fossils are similar to those described by Lillegraven (1969). Many of these specimens may eventually be referred to A. jasoni or A. marshi (see discussion above). Dental measurements are given in Table 25.
Isolated lower molars of smaller species of Alphadon are difficult to separate with confidence and are distinguished by differences in size and proportions (Clemens, 1966, 1973; Lillegraven, 1969), but there is some degree of overlap in their ranges of variation. Specimens from McGuire Creek referred to A. "wilsoni" are similar in size and proportions to the teeth described by Lillegraven (1969) and can be separated from A. marshi by these criteria (but see discussion above). Dental measurements are given in Table 26.
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Alphadon marshi Simpson, 1927b
Alphadon marshi Simpson, 1927b, p. 125
Type . YPM 13659, right M3/ (Simpson, 1929a, fig. 45F).
Type locality . Lance Formation, Niobrara County, Wyoming.
Referred specimen . One /M4, UCMP 134777, from loc. V87072.
Locality . UCMP loc. V87072.
Distribution . Possibly Fruitland and Kirtland formations, New Mexico (Edmontonian); Lance Formation, Wyoming, and Scollard Formation, Alberta (both Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Revised diagnosis . Clemens, 1966, p. 5, and Lillegraven, 1969, p. 33.
Description . A single specimen is referred to A. marshi on the basis of size and the proportions of length to trigonid and talonid width as described by Lillegraven (1969). Measurements of UCMP 134777 are: L: 2,38; W/Tri: 1.47; W/Tal: 1.27; L/W-Tri: 1.62; L/W-Tal: 1.87.
Alphadon rhaister Clemens, 1966
Table 27
Alphadon rhaister Clemens, 1966, p. 11.
Type . UCMP 50292, fragmentary left maxilla with M1-3/.
Type locality . UCMP loc. V5815, Lance Formation, Wyoming.
Referred specimens . One /Mx talonid from loc. V87036. One /Mx trigonid from loc. V87052. One /M2 or /M3, from loc. V87072. One /Mx from loc. V87074. Two /M1's, from loc. V87098.
Localities . UCMP locs. V87036, V87052, V87072, V87074, and V87098.
Distribution . Possibly St. Mary River Formation, Alberta (Edmontonian); Lance Formation, Wyoming, and Scollard Formation, Alberta (both Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Description . Fossils from McGuire Creek referred to Alphadon rhaister are similar to those described by Clemens (1966) from the type Lance Formation. The positioning of the intersection of the trigonid and the cristid obliqua below the protoconid-metaconid
| ||||||||||||||||||||||||||||||||||||||||||||||||
notch separates them from teeth of similar-sized species of Pediomys . In size (Table 27) they are consistently within the range of variation exhibited by Alphadon rhaister (Clemens, 1966, table 19). Morphologically, they add no new information to the original description of the species.
Alphadon sp indet.
Table 28
Referred specimens . One M2/ from UCMP loc. V87035. One /M1 from UCMP loc. V87037. One M4/ from UCMP loc. V87072. One /M1 from UCMP loc. V87098. Five fragmentary molars from various UCMP localities.
Localities . UCMP locs. V87035, V87037, V87038, V87072, V87074, and V87098.
Description . The four specimens listed in Table 28 are intermediate in size and proportions between those of Alphadon marshi and A. "wilsoni" . Recently, Johnston and Fox (1984) and Fox (1989) described isolated molars of Alphadon from the Long Fall (Ravenscrag Formation) and Wounded Knee (Frenchman Formation) localities in Saskatchewan which are also intermediate in size between A. "wilsoni" and A. marshi . Apparently these fossils belonged to an as then undescribed species of the genus (referred to as Alphadon sp.) which was to be named and characterized when a larger sample was collected (Johnston and Fox, 1984; Fox, 1989). Subsequently, Storer (1991) referred all Alphadon specimens from Wounded Knee to a new species, A. jasoni .
McGuire Creek fossils of Alphadon within the size range that Johnston and Fox (1984) and Fox (1989) assigned to the unnamed species may be conspecific with A. jasoni or A. marshi . Pending formal publication of the revision of species of Alphadon in progress by Case and Johanson (described above), the four McGuire Creek specimens listed in Table 28 are referred to Alphadon sp. indeterminate.
The other five specimens included here are small but are too fragmentary to be referable to any of the small species of Alphadon with confidence.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Subfamily PERADECTINAE (Crochet, 1979) Reig, 1981
Peradectes Matthew and Granger, 1921
Peradectes cf. P. pusillus Matthew and Granger, 1921
Table 29
Thylacodon pusillus Matthew and Granger, 1921, p. 2.
Type . AMNH 16414, dentary fragment with M2-3, M1 talonid (Matthew, 1937, fig. 82).
Type locality . " . . . upper level of the Puerco Formation near Ojo Alamo, San Juan Basin, New Mexico . . . " (Matthew and Granger, 1921, p. 2).
Referred specimens . Associated M1/, UCMP 132299, and M2/, UCMP 132300, from loc. V84194. One /M1 from loc. V87035. One /M2 from loc. V87038. One M3/, 1 /M2 from loc. V87037. One M1/, 1 /M3 from loc. V87072. One M1/ from loc. V87074.
Localities . UCMP locs. V84194, V87035, V87037, V87038, V87072, and V87074.
Distribution . Upper Hell Creek and lower Tullock formations, Montana (both Puercan).
Comments . Reference of this material to Peradectes cf. P. pusillus is based on comparison with material from the Hell's Hollow Local Fauna (UCMP locs. V74110 and V74111) described by Archibald (1982). Archibald's (1982) referral of the Hell's Hollow material to Peradectes cf. P. pusillus was based on an unpublished revision of North American didelphids in progress by W. A. Clemens at that time, which included the referral of Thylacodon pusillus to the genus Peradectes . Alternatively, Krishtalka and Stucky (1983) have argued that Thylacodon pusillus may not be a species of Peradectes . Clemens has now included the Hell's Hollow material in his study and the manuscript is presently in the final stages of preparation (pers. comm., Clemens, 1989), publication of which should aid in settling this question.
Description. Peradectes cf. P. pusillus was adequately described by Archibald (1982). With the exception of one specimen, all teeth referred to this species from McGuire Creek closely resemble those from Hell's Hollow in size and morphology (Table 29). An isolated M1/, UCMP 134779 from V87072, differs in the morphology of the C region of the stylar shelf. All M1/'s from Hell's Hollow have a large B cusp and slightly smaller
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
and subequal C and D cusps (Archibald, 1982). These 3 stylar cusps are well separated. UCMP 134779 has 2 stylar cusps in the region normally occupied by a single C cusp in the Hell's Hollow specimens. The anterior of the two is very small and is not well separated from cusp B. It is situated on the labial edge of the stylar shelf anterior to a slight indentation on the midline of the shelf, which is suggestive of a weakly developed ectoflexus. The posterior C cusp is larger, but still much smaller and well separated from cusp D and the other more anteriorly positioned C cusp. The posterior C cusp is shortened in an antero-posterior orientation, giving the cusp a sharp ridge-like appearance. Otherwise, UCMP 134779 is very similar to M1/'s from Hell's Hollow.
Order SPRASSODONTA (Ameghino, 1894) Marshall, Case, and Woodburne, 1990
Superfamily BORHYAENOIDEA (Ameghino, 1894) Simpson, 1930
Family STAGODONTIDAE Marsh, 1889b
Didelphodon Marsh, 1889a
Didelphodon vorax Marsh, 1889a
Table 30
Didelphodon vorax Marsh, 1889a, p. 88 (See Clemens, 1966, p. 60, for synonymies).
Type . YPM 11827, left M2/ (Marsh, 1889a, pl. IV, figs. 1-3).
Type locality . Mammal locality no. 1 of Lull (1915), UCMP loc. V5003, Lance Formation, Wyoming.
Referred specimens . Left maxillary fragment with complete or partial alveoli for C/, and P1-3/, UCMP 134795; left edentulous dentary fragment, UCMP 134793; maxillary fragment with labial alveoli of Mx, UCMP 134794; 1 /M4 fragment, 1 /P1 fragment, 1 /P1, 1 Mx/ fragment, 1 P3/, from loc. V85092. One P2/ from loc. V87029. One /P3, 1 M1/ from loc. V87034. One /Px fragment, 1 /M4 talonid from loc. V87035. One P3/
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
fragment from loc. V87037. One /M2 from loc. V87070. One /DP3 from loc. V87072. One Mx/ fragment from loc. V87073. One Mx/ fragment from loc. V87078. Left edentulous dentary fragment, UCMP 132807, from loc. V87083. One DP3/ fragment from loc. V87098. One /Mx fragment from loc. V87151.
Localities . UCMP locs. V85092, V87029, V87034, V87035, V87037, V87070, V87072, V87073, V87078, V87083, V87098, and V87151.
Distribution . Lance Formation, Wyoming, Scollard Formation, Alberta, and Hell Creek Formation, South Dakota (all Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Revised diagnosis . Clemens, 1966, p. 60, with modifications by Clemens, 1968, pp. 4-10.
Description . Isolated teeth from McGuire Creek referred to Didelphodon vorax are similar to those from the Lance Formation, Wyoming, described by Clemens (1966), with one exception. UCMP 134563, a /P1, is larger (Table 30) than those from the Lance Formation (Clemens, 1966, table 13), but with regard to the range in variation of premolar size exhibited by the species this is not considered significant. Dental measurements are given in Table 30.
Discussion . The dental arcade of stagodontid marsupials is notable for the presence of bulbous premolars, each with a massive main cusp and a large accessory lobe. Configuration of the upper premolars of Didelphodon and other stagodontid marsupials was uncertain because of the lack of specimens preserving this portion of the skull. Isolated upper premolars of Didelphodon were interpreted to be oriented with their accessory lobes on either their labial (Clemens, 1966) or lingual (Clemens, 1968; Lillegraven, 1969; Archibald, 1982; Fox and Naylor, 1986) sides.
Lofgren (1992) presented the upper premolar orientation of Didelphodon vorax using an edentulous maxillary fragment (UCMP 134795) from the upper Hell Creek Formation at McGuire Creek (UCMP loc. V85092). The maxillary fragment contains a number of complete or partial alveoli, into which isolated premolars of D. vorax from the type Lance Formation, Wyoming, were placed. This process indicates that the accessory lobes are lingual on P2/ -P3/ and only a single alveolus is available for P1/ (Lofgren, 1992). Isolated premolars identified as P1/'s in previous studies have two roots (Clemens, 1966; Lillegraven, 1969), but these teeth may actually be P2/'s (Lofgren, 1992). In any case, the only definitive statement concerning P1/ morphology is provided by UCMP 134795, which has but a single alveolus available for P1/.
Cohort AUSTRALIDELPHIA Szalay, 1982
Order MICROBIOTHERIA Ameghino, 1889
Superfamily MICROBIOTHERIODEA (Ameghino, 1887) Reig, Kirsch, and Marshall, 1985
Family PEDIOMYIDAE (Simpson, 1927a) Clemens, 1966
Pediomys Marsh, 1889a
A systematic revision of Pediomys is presently underway by R. Cifelli. These revisions are not available at present, but apparently changes in generic reference are forthcoming.
Pediomys krejcii Clemens, 1966
Table 31
Pediomys krejcii Clemens, 1966, p. 43.
Type . UCMP 51390, fragmentary left maxilla with P3/, M1-3/ (Clemens, 1966, fig. 26).
Type locality . UCMP loc. V5711, Lance Formation, Wyoming.
Referred specimens . Right dentary fragment with M4, UCMP 132490, from loc. V87036. Two Mx/'s, 1/Mx, from loc. V87037. One /M2 from loc.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
V87072. One /M3 from loc. V87074. One M1/, 1 /M1 trigonid, both from loc. V87151.
Localities . UCMP locs. V87036, V87037, V87072, V87074, and V87151.
Distribution . Possibly St. Mary River Formation, Alberta (Edmontonian); Lance Formation, Wyoming, and Scollard Formation, Alberta (both Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Description . Reference of the upper molars listed in Table 31 to Pediomys krejcii is based on their small size and the absence of a stylar cusp C on UCMP 132176. P. krejcii is distinguished from P. cooki by its small size, and can be separated from P. elegans by the absence of stylar cusp C. UCMP 132176 differs slightly from the original description of the species given by Clemens (1966) in that it has a distinct anterolingual cingulum. Lillegraven (1969) noted the presence of this character in the P. krejcii sample from the upper Edmonton Formation of Alberta, but considered it to have little taxonomic significance because of the highly variable development of the cingula.
Specimens of the lower dentition from McGuire Creek referred to P. krejcii are similar to those from the type Lance Formation described by Clemens (1966). Dental measurements are presented in Table 31.
Pediomys elegans Marsh, 1889a
Tables 32-33
Pediomys elegans Marsh, 1889a, p. 89.
Type . YPM 11866, broken and heavily weathered upper molar (Marsh, 1889a, pl. IV, figs. 23-25).
Neotype . CM 11658, fragmentary right maxilla with M2-4, designated as neotype by Simpson (1929b, p. 111).
Type locality . Mammal locality no. 1 of Lull (1915) = UCMP loc. V5003. Neotype from somewhere in the type Lance Formation, probably loc. V5003.
Referred specimens . One M2/ from loc. V87028. One M1/ from loc. V87037. One /M1, 1 /M2, from loc. V87038. One /M3, 1 M3/, from loc. V87051. One /M4 from loc.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
V87052. Two /M3's, 1 /M4, from loc. V87071. Left dentary fragment with /M1, /M3-4, UCMP 134773, left dentary fragment with /M3-4, UCMP 133462, 2 M1/'s, 2 /M1's, 2 /M2's, 1 /M4, 2 /Mx's, all from loc. V87072. Two M3/'s, 1 /M3, 1 /Mx, from loc. V87074. One M2/ from loc. V87098.
Localities . UCMP locs. V87028, V87037, V87038, V87051, V87052, V87071, V87072, V87074, and V87098.
Distribution . Lance Formation, Wyoming, and Scollard Formation, Alberta (both Lancian); Frenchman Formation, Saskatchewan (Lancian, in part); Hell Creek Formation, Montana (Lancian and *Puercan); Ravenscrag Formation, Saskatchewan (*Puercan).
Revised diagnosis . Clemens, 1966, p. 35.
Description . Upper molars referred to P. elegans agree closely in size (Table 32) and stylar cusp morphology to Clemens' (1966) description of the species. A large or distinct stylar cusp C is evident on all specimens. This feature separates P. elegans from the similar-sized species of Pediomys, P. cooki (Clemens, 1966). Also, the McGuire Creek fossils tend to have a large D cusp which is usually small in P. cooki (Clemens, 1966).
Dental measurements of lower molars of P. elegans are presented in Table 33. Lower molars of P. elegans and P. cooki are distinguished primarily by size (Clemens, 1966). The specimens from McGuire Creek consistently fall into the size range displayed by P. elegans (Clemens, 1966, table 22). However, there is some degree of overlap in size between the two species and some fossils referred to P. elegans may actually
belong to P. cooki . Although possible, this is considered unlikely, because all of the upper molars in this size range from McGuire Creek are clearly referable to P. elegans , and most of these come from the same localities as the lower molars.
Morphologically, the lower molars of P. elegans from McGuire Creek are similar to Clemens' (1966) description of the species, and only one addition is necessary. All /M3's and /M4's and two-thirds of the other molars (90% of total sample, n=21) have a small entoconulid. In most cases, the entoconid is larger and higher than both the entoconulid and the hypoconulid. In size and height, the hypoconulid and entoconulid are subequal and the entoconulid is usually not well separated from the entoconid except in /M3's. A survey of the P. elegans sample from the type Lance Formation of Wyoming (UCMP loc. V5620) shows that only 30% of the lower molars (n=40) have an entoconulid, and the majority of these are /M4's. The sample of P. elegans from the Edmonton Formation of Alberta is small, but the only lower molar (an /M2) figured by Lillegraven (1969, figure 22) has this cusp. The variation in this character probably is not of taxonomic significance.
Pediomys hatcheri (Osborn, 1898) Simpson, 1927b
Table 34
Protolambda hatcheri Osborn, 1898, p. 172 (see Clemens, 1966, p. 45, for synonymies).
Type . AMNH 2202, fragmentary right maxilla with M3/, designated as a lectotype by Clemens, 1966, p. 45 (Osborn, 1898, fig. 1A, center); AMNH 2203, isolated M2/, designated as a syntype by Clemens, 1966, p. 45.
Type locality . "Lance Creek area, Lance Formation, Wyoming" (Clemens, 1966, p. 45).
Referred specimens . One /Mx from loc. V87035. Left dentary fragment with /M3-4, UCMP 132599, from loc. V87037. One M2/, 1 /Mx trigonid, both from loc. V87072. One /M2 from loc. V87098. One M2/ from loc. V87151.
Localities . UCMP locs. V87035, V87037, V87072, V87098, and V87151.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distribution . Lance Formation, Wyoming, Scollard Formation, Alberta, and North Horn Formation, Utah (all Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Revised diagnosis . Clemens, 1966, p. 45.
Description . In both the M2/'s referred to Pediomys hatcheri , the stylar shelf is continuous across the paracone, the B cusps are absent, and the C cusps are small. In these respects and in size (Table 34), they agree closely with the original description of the species by Clemens (1966). One small difference is noteworthy; UCMP 132178 has a small anterolingual cingulum.
Lower molars of Pediomys hatcheri from McGuire Creek agree with Clemens' (1966) original descriptions, and only one addition is necessary. The dentary fragment, UCMP 132599, has a long narrow talonid on /M4. Consequently, the length of this tooth falls outside the range of variation observed in the sample from the type Lance Formation.
Pediomys florencae Clemens, 1966
Table 35
Pediomys florencae Clemens, 1966, p. 50.
Type . UCMP 51440, fragmentary left maxilla with M2-3 (Clemens, 1966, fig. 31).
Type locality . UCMP loc. V5820, Lance Formation, Wyoming.
Referred specimens . Right edentulous dentary fragment, UCMP 134792, from loc. V85092. One M1/, 1 M4/, from loc. V87035. One /Mx trigonid from loc. V87038. One /M3 from loc. V87040. One M1/ from loc. V87072. One Mx/ fragment, 1 /M2, both from loc. V87151.
Localities . UCMP locs. V85092, V87035, V87038, V87040, V87072, and V87151.
Distribution . Lance Formation, Wyoming (Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Description. Pediomys florencae is the largest species of the genus, and the fragmentary specimens UCMP 134792 and UCMP 134789 are referred to this taxon primarily on this basis. A large edentulous dentary fragment, UCMP 134792, is also referred to
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
the species. It compares closely to mandibles of P. florencae from UCMP locality V5620 from the type Lance Formation.
Two complete lower molars from McGuire Creek are referred to P. florencae . One of these, an /M3 (UCMP 133153), is larger than the single /M3 from the type Lance Formation described by Clemens (1966). Until further sampling proves otherwise, this size difference is attributed to variation within the species.
Of the three upper molars referred to P. florencae , two provide some additional morphological data. A well preserved M1/, UCMP 132517, has small C and D stylar cusps, and cusp B is absent. Also, the stylar shelf is not continuous across the paracone. On UCMP 132515, an M4/, the stylar shelf extends to a small indentation posterior to a small C cusp, cusp B is absent, and a large parastyle is present. The metacone is much smaller than the paracone and is near the posterolabial edge of the tooth. A weak anterolingual cingulum and both conules are present. Dental measurements are provided in Table 35.
Pediomys sp. indet.
Referred specimens . Two /Mx talonids from loc. V87037. One /Mx talonid from loc. V87038. One M2/?, UCMP 133181, from loc. V87071.
Localities . UCMP locs. V87037, V87038, and V87071.
Comments . On UCMP 133181, the stylar shelf labial to the paracone is present but weakly developed, and cusp B is absent. Unfortunately, the posterolabial quarter of the tooth, including the area of the stylar shelf which would contain cusps C and D (if present), is missing. On the basis of size, this specimen could be referred to P. cooki or P. elegans , but loss of the posterior part of the stylar shelf prevents reference to one or the other species.
Three lower molar talonids are referred to Pediomys sp. indeterminate, because they are too fragmentary for species identification.
Infraclass EUTHERIA (Gill, 1872) Huxley, 1880
Cohort EPITHERIA McKenna, 1975
Superorder INSECTIVORA (Illiger, 1811) Novacek, 1986
Order LEPTICTIDA (McKenna, 1975) Novacek, 1986
Family GYPSONICtopIDAE
Gypsonictops Simpson, 1927a
Gypsonictops illuminatus Lillegraven, 1969
Tables 36-37; Figure 23
Gypsonictops illuminatus Lillegraven, 1969, p. 51.
Type . UA 2447, right maxillary fragment with P3-4/, M1-3/ (Lillegraven, 1969, fig. 27-6).
Type locality . Loc. KUA-1, Scollard Formation, Alberta.
Referred specimens . One P4/ fragment from loc. V85092. One /Mx, 1 Mx/, from loc. V87035. One /M1, 2 DP4/'s, 1 /DP4, from loc. V87038. One /M2, 1 M3/, from loc. V87051. Two /M1's from loc. V87071. One /P4, 2 /M3's, 1 M1/, 1 Mx/, from loc.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
V87072. One /M1, 1 /M2, from loc. V87074. One /DP4, 1 P3/, from loc. V87098. One M3/ from loc. V87101. 1 M2/, 1 P4/, from loc. V87151. One Mx/ fragment from loc. V87153.
Localities . UCMP locs. V85092, V87035, V87038, V87051, V87071, V87072, V87074, V87098, V87101, V87151, and V87153.
Distribution . Scollard Formation, Alberta (Lancian); possibly Frenchman Formation, Saskatchewan (Lancian, in part); Hell Creek Formation, Montana (Lancian and *Puercan); Ravenscrag Formation, Saskatchewan (*Puercan).
Description . Lower molars from McGuire Creek referred to Gypsonictops illuminatus are consistently within the size range of G. illuminatus from the Scollard Formation of Alberta described by Lillegraven (1969). The dentition of G. hypoconus , a smaller species known from the Lance Formation of Wyoming (Clemens, 1973), is significantly smaller. The other character that differentiates the lower molars of these two species is the relative height of the trigonid: G. illuminatus has proportionately lower trigonids than G. hypoconus (Lillegraven, 1969; Clemens, 1973). The /M1's and /M2's are identified by the method outlined by Clemens (1973). A single /DP4 (UCMP 133079) is referred to G. illuminatus , on the basis of size and comparison to /DP4's of G. illuminatus described by Lillegraven (1969, figure 27, 2a-c). Dental measurements are given in Table 36.
Upper molars and premolars from McGuire Creek also consistently fall within the range of variation in size displayed by G. illuminatus and are significantly larger than
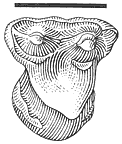
Figure 23
Gypsonictops illuminatus
Lillegraven. Isolated
right DP4/, UCMP 133077,
locality V87038, occlusal
view. Scale bar = 2 mm.
those of G. hypoconus . Also, unworn upper molars from McGuire Creek are similar to G. illuminatus in the development of an anterior recumbency in the protocone. Three DP4/'s from McGuire Creek are referred to G. illuminatus . Previously, DP4/'s of G. illuminatus were unknown. Fossils from McGuire Creek are identical to DP4/'s of G. hypoconus described by Clemens (1973) from the Lance Formation, but are significantly larger and have an anteriorly recumbent protocone (Figure 23). Dental measurements are given in Table 37.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Infraclass EUTHERIA, incertae sedis
Family PALAEORYCITIDAE (Winge, 1917) Simpson, 1931
Batodon Marsh, 1892
Batodon tenuis Marsh 1892
Table 38
Batodon tenuis Marsh, 1892, p. 258.
Type . USNM 2139, fragmentary left dentary with part of the canine, alveolus of /P1, and /P2-4 (Marsh, 1892, pl. XI, fig. 5; Simpson, 1929a, fig. 55; Clemens, 1973, fig. 25d and e).
Type locality . Mammal locality no. 1 of Lull (1915); UCMP loc. V5003, Lance Formation, Wyoming.
Referred specimens . One /P4, 1 M2/ from loc. V87038. One /M2 from loc. V87074. One /M2 from loc. V87151.
Localities . UCMP locs. V87038, V87074, and V87151.
Distribution . Lance Formation, Wyoming, and Scollard Formation, Alberta (both Lancian); Hell Creek Formation, Montana (Lancian and *Puercan).
Revised Diagnosis . Clemens, 1973, p. 67-68.
Description . The sample of Batodon tenuis from McGuire Creek is small, consisting of four teeth from three widely scattered localities. Batodon tenuis has been adequately described by Lillegraven (1969) and Clemens (1973), and the specimens from McGuire Creek yield little additional morphological information. Dental measurements are provided in Table 38.
| ||||||||||||||||||||||||||||||||||||||||||
Procerberus Sloan and Van Valen, 1965
Procerberus formicarum Sloan and Van Valen, 1965
Table 39
Procerberus formicarum Sloan and Van Valen, 1965, p. 225.
Type . UMVP 1460, dentary fragment with /P3-4, /M1 (not figured, but isolated teeth figured, Sloan and Van Valen, 1965, fig. 6; see also Lillegraven, 1969, fig. 32-1,2,3, and Van Valen, 1969, fig. 3).
Type locality . Bug Creek Anthills, Hell Creek Formation, Montana.
Referred specimens . One M3/ from loc. V84193. One M1/ from loc. V84194. One /M2, 1 M2/ from loc. V87028. Two /P4's from loc. V87030. One /M1, 1 /M2, 2 /M3's, 1 P3/, 1 P4/ from loc. V87035. One /M1 from loc. V87036. Two /P4's, 1 /M1, 1 /M2, 1 /M3, 1 P3/, 2 P4/'s, 2 M1/'s, 2 M2/'s, 1 M3/, 2 Mx/'s from loc. V87037. Four /P4's, 3 /M1's, 4 /M2's, 4 /M3's, 1 DP4?/, 2 P4/'s, 2 M1/'s, 3 M2/'s, 2 M3/'s from loc. V87038. One /P4, 2 /M1's, 1 P3/, 1 M1/ from loc. V87051. One /P4, 1 M3/ from loc. V87071. One /P3, 3 /P4's, 6 /M1's, 2 /M2's, 2 /M3's, 1 DP4/, 3 P3/'s, 4 P4/'s, 10 M2/'s, 2 M3/'s from loc. V87072. Two M1/s from loc. V87073. Four /M1's, 1 /M2, 1 /M3, 1 P3/, 5 P4/'s, 2 M1/'s, 2 M2/'s, 2 M3/'s, 3 Mx/'s from loc. V87074. Two /P4's, 1 /M1, 1 /M2, 1 /M3, 1 P4, 2 M1/'s, 1 M2/, 1 Mx/ from loc. V87098. One P4/ from loc. V87114. One /DP4 trigonid, 3 /M1's, 2 /M2's, 4 /M3's, 2 P4/'s, 2 M1/'s, 6 M2/'s, 1 M3/, 1 Mx/ from loc. V87151. One /M3 from loc. V87153.
Localities . UCMP locs. V84193, V84194, V87028, V87030, V87035, V87036, V87037, V87038, V87051, V87071, V87072, V87073, V87074, V87098, V87114, V87151, and V87153.
Distribution . Upper Hell Creek and lower Tullock Formations, Montana (both Puercan); possibly Ravenscrag Formation, Saskatchewan (Puercan).
Description. Procerberus formicarum from the upper Hell Creek Formation at McGuire Creek is similar in size and morphology to fossils from Bug Creek Anthills, the type locality (Table 39). The dental morphology of P. formicarum has been described in detail by Sloan and Van Valen (1965) and Lillegraven (1969), and additional comments will be restricted to the DP4/, which is not described in those works.
The only DP4/ (UCMP 134706) that can be confidently referred to P. formicarum is unworn. The crown is nearly equal in length and width (L: 2.56; W-A: 2.56; W-P: 2.61), giving the tooth a triangular shape. In gross morphology, the DP4/ of P. formicarum resembles those referred to Cimolestes by Lillegraven (1969) and Clemens (1973) but differs significantly in the following ways: The paracone and metacone are less conate, and the metacrista and paracrista are less distinct, the stylar shelf is more reduced, with the stylar area labial to the paracone markedly so, the conules are less distinct and lack internal wings, and short and weakly developed lingual cingula are present.
Superorder UNGULATA (Linnaeus, 1766) Novacek, 1986
Order "CONDYLARTHRA"
(Quotation marks indicate a paraphyletic taxon; Prothero et al., 1988.)
Most of the mammalian taxa found in K-T transition local faunas in eastern Montana (upper Hell Creek and lower Tullock formations) have been adequately described in previous works (e.g., Clemens, 1964, 1966, 1973; Sloan and Van Valen, 1965; Lillegraven, 1969; Archibald, 1982). "Condylarths" are a notable exception, because the description of many taxa consists of only one or two sentences given with their original diagnoses (Sloan and Van Valen, 1965; Van Valen, 1978). Archibald (1982) provided more complete descriptions of Oxyprimus erikseni, Mimatuta morgoth , and Mimatuta minuial . Recently, Luo (1989, 1991) gave detailed descriptions of Protungulatum donnae, Mimatuta morgoth , and Oxyprimus erikseni . However, Protungulatum gorgun and
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Ragnarok nordicum are still inadequately described and poorly known. A more complete description of the last two species based on new material from McGuire Creek, is given below. Oxyprimus erikseni, Mimatuta morgoth, Mimatuta minuial , and Protungulatum donnae also are present at McGuire Creek, and additions to their descriptions are given where deemed necessary.
During systematic analysis of the "condylarths" from McGuire Creek, it became apparent that many of the "condylarths" from Mantua Lentil, Polecat Bench Formation (Fort Union Formation), Wyoming, are similar to those from the upper Hell Creek Formation of Montana. Because regional biostratigraphic and biochronological correlations during the Late Cretaceous?-Early Tertiary are based in part on species-level differences within "condylarth" lineages, it became necessary to evaluate the validity of species of genera (Oxyprimus, Ragnarok , and Mimatuta ) that occur in both the Mantua Lentil and Hell Creek areas. Epoxy casts of Oxyprimus putorius, O. galadrielae, Ragnarok nordicum , and Mimatuta minuial from Mantua Lentil were available for study. Descriptions of these taxa and comparisons to McGuire Creek "condylarths" are given where appropriate.
Family ARCTOCYONIDAE (Giebel, 1855) Murray, 1866
Subfamily OXYCLAENINAE (Scott, 1892) Matthew, 1937
Protungulatum Sloan and Van Valen, 1965
Protungulatum donnae Sloan and Valen, 1965
Tables 40-42, 54
Protungulatum donnae Sloan and Van Valen, 1965, p. 226.
Type . SPSM 62-2028, left mandible with /P2-4, /M1-3 (Sloan and Van Valen, 1965, fig. 7).
Type locality . Bug Creek Anthills, Hell Creek Formation, Montana.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Referred specimens . Dentary with /M2-3, UCMP 132461, from loc. V86031. Dentary with /M2-3, UCMP 132341; 1 /M3 from loc. V87029. Dentary with /M2, UCMP 132440, from loc. V87031. One M1/, 1 /M2 from loc. V87034. Dentary with /M2-3, UCMP 132498; dentary with /M1-2, UCMP 132499; dentary with /M2, UCMP 132497, from loc. V87035. One M1/ from loc. V87036. Dentary with /P4-/M1, UCMP 132595; 2 /M1's, 1 M3/ from loc. V87037. One M1/ from loc. V87038. One /M2 from loc. V87040. Dentary with /M2-3, UCMP 133517; maxilla with M2-3, UCMP 134696; 1 M2/ from loc. V87072. One /M3 from loc. V87082. One M2/ from loc. V87084. Dentary with /M1 talonid, /M2, /M3 trigonid, UCMP 133838; 2 /M1's, 1 /M2, 1 /M3 from loc. V87098. Maxilla with M1-2/, UCMP 133820; 1 /M2 from loc. V87101. One /M2 from loc. V87108. Two /M1's, 1 /M2, 1 M3/ from loc. V87151. One /M2 from loc. V88038.
Localities . UCMP locs. V86031, V87029, V87031, V87034, V87035, V87036, V87037, V87038, V87040, V87072, V87082, V87084, V87098, V87101, V87108, V87151, and V88038.
Distribution . Upper Hell Creek Formation, Montana (Puercan); possibly lower Tullock Formation, Montana (Puercan); possibly upper Frenchman Formation and Ravenscrag Formation, Saskatchewan (both Puercan).
Description . McGuire Creek fossils referable to Protungulatum can be separated into
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
two species of similar morphology but of significantly different size ranges. The larger species is the same size as P. gorgun from Harbicht Hill, Hell Creek Formation, described by Van Valen (1978). The smaller of the two species is referred to P. donnae from Bug Creek Anthills, Hell Creek Formation, described by Sloan and Van Valen (1965).
In addition to the original description, the dentition of Protungulatum donnae was described in further detail and contrasted with other Late Cretaceous?-early Paleocene "condylarths" (e.g. Mimatuta, Oxyprimus , and Ragnarok ) by Archibald (1982), Middleton (1983), and Luo (1989, 1991). Therefore, only a few comments are required concerning additional morphological data from McGuire Creek.
Upper molars of Protungulatum donnae are similar in size to those of Mimatuta , but can be separated from the latter by their steeper and shorter lingual protoconal slopes (Archibald, 1982; Luo, 1989, 1991). This is expressed by lower A/W-A ratios for Protungulatum donnae (Tables 40, 54) than for Mimatuta morgoth (Tables 53, 54) at McGuire Creek. Lower molars of Mimatuta are also similar in size to those of Protungulatum donnae (Tables 41, 42 and 55, 56), but are distinguished by the labial shift of the paraconids in the former (Archibald, 1982). This shift is accompanied by a deepening of the valley between the paraconid and metaconid (Luo, 1989). Also, lower molars of M. morgoth have on average slightly wider talonids than P. donnae (Archibald, 1982).
Upper and lower molars of Protungulatum donnae and P. gorgun from McGuire Creek are similar, but differ significantly in size (Tables 40, 41, 42 and 43, 44, 45). The size separation is great, with minimal overlap evident, and presumably denotes the presence of two separate species of Protungulatum . Lower molars of P. donnae also can be distin-
guished from those of P. gorgun by the development of a large anteriorly projecting paraconid in the latter (Van Valen, 1978).
A taxonomic problem is evident when comparing the McGuire Creek sample of Protungulatum donnae to the type and only specimen of Protungulatum mckeeveri (/P4-/M3, UCMP 121782). P. mckeeveri was distinguished from P. donnae by more reduced paraconids on /M1-3, longer /P4 and /M1-2 relative to width, and other minor differences in the proportions of the lower molars (Archibald, 1982). In contrast to the Bug Creek Anthills sample, the McGuire Creek sample referred to P. donnae has a range of variation in size that completely includes that of P. mckeeveri (Table 41). The McGuire Creek sample of P. donnae comes from a number of different localities in the upper Hell Creek Formation that probably differ slightly in age. Therefore, these two species may be present in the upper Hell Creek Formation at McGuire Creek, but are present at localities of slightly differing age. Alternatively, these species may have coexisted and were present at a single site. It is also possible that P. donnae and P. mckeeveri are synonymous, and data presented below suggest that this is probably true. A dentary fragment (UCMP 132595) here referred to P. donnae contains a /P4 of the size and proportions appropriate for P. donnae and an /M1 of the size and proportions of /M1's of P. mckeeveri (Tables 41, 42). The few isolated molars collected from the same site as UCMP 132595 are referable to P. donnae .
Another locality, V87098, produced a dentary fragment (UCMP 133838, Table 42) of Protungulatum donnae along with isolated molars that could be referred to P. mckeeveri . Other than slight differences in size and proportion, the only character that separates the two species is the slightly more reduced paraconids of P. mckeeveri (Archibald, 1982). A brief survey of the UCMP collections from Bug Creek Anthills, the type locality of P. donnae , shows that a dentary fragment (UCMP 105006) has more reduced paraconids than P. mckeeveri . Given the data presented above, I suspect that P. donnae and P. mckeeveri are synonymous. However, a comparative study of P. mckeeveri and a large sample of "condylarths" from Bug Creek Anthills is needed to settle the matter.
Protungulatum gorgun Van Valen, 1978
Tables 43-45; Figures 24-26
Protungulatum gorgun Van Valen, 1978, p. 52.
Type . AMNH 35987, right mandible with /M2 (Van Valen, 1978, pl. 1, fig. 4).
Type locality . Harbicht Hill, Hell Creek Formation, Montana.
Referred specimens . Associated teeth /P4-/M3 (evidently from a dentary that was destroyed during screenwashing), UCMP 134622; dentary with /M3, UCMP 132318, from loc. V87028. One M1/, 1 M2/, 1 /M1 from loc. V87029. One /M2 from loc. V87034. One M1/, 2 M2/'s from loc. V87035. Dentary with /P2-3, /M1-2, UCMP 134558; 1 /M2 from loc. V87037. Dentary with /M2, UCMP 133064; 1 /M2, 1 M2/ from loc. V87038. One /M2, 1 M3/ from loc. V87049. Dentary with /M2, UCMP 133126, from loc. V87067. One M2/ from loc. V87071. One M2/ from loc. V87077. Dentary with /M1-2, UCMP 132806, from loc. V87084. One /M2 from loc. V87088. One M3/, 1 M2/ from loc. V87098. Dentary with /M2-3, UCMP 133817, from loc. V87107.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Dentary with /M1-2, UCMP 133837, from loc. V87115. One M2/ from loc. V87119. Dentary with /M2-3, UCMP 132436, from loc. V87124.
Localities . UCMP locs. V87028, V87029, V87034, V87035, V87037, V87038, V87049, V87067, V87071, V87077, V87084, V87088, V87098, V87107, V87115, V87119, and V87124.
Distribution . Upper Hell Creek Formation, Montana (Puercan).
Description . Of all the "condylarths" known to occur in the upper Hell Creek Formation in eastern Montana, Protungulatum gorgun is undoubtedly the poorest known. Van Valen's (1978) diagnosis of P. gorgun is brief, and he provides little additional morphological description. The only other description of P. gorgun was provided by Lupton et al. (1980), who described two dentary fragments collected from Chris' Bonebed, a Milwaukee Public Museum locality in the upper Hell Creek Formation in McCone County. The upper dentition of P. gorgun has never been described.
Fossils from McGuire Creek are referred to Protungulatum gorgun on the basis of comparisons to an epoxy cast of the type (AMNH 35987), an isolated /M2 from Harbicht Hill. A number of dentaries from McGuire Creek referable to P. gorgun contain the /M2 with other parts of the lower dentition (Table 43). These fossils form the basis for the description of the lower dentition presented below.
The portion of the mandible anterior to /P4 of Protungulatum gorgun is preserved only in UCMP 134558 (Figure 24). UCMP 134558 has two mental foramina, one below the /P1-/P2 diastema, and a second below the posterior root of /P3. Only the root of
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
the canine is preserved, and it is larger than the roots of /P1 and /P2. The canine root is recurved under the root of /P1. /P1 is missing, but its alveolus is angled posteriorly, indicating that /P1 was slightly procumbent. There is a short diastema between /P1-/P2 and one nearly twice as long between /P2-/P3. The remaining premolars have two roots. /P2 has a large central cusp with weakly developed anterior and posterior midline ridges. Also present is a small posterior basal cusp, but an anterior basal cusp is lacking. /P3 has a large central cusp that is higher and more robust than that of /P2. A small anterior basal cusp is present and is shifted lingual to the midline of the tooth. The posterior portion of the /P3 is missing because of breakage.
Only one dentary (UCMP 134622) has a /P4 (Figure 25D-F), and it is moderately worn. The /P4 has a tall, robust protoconid with a small metaconid placed low on its side. The paraconid is large, slightly procumbent, well separated from the protoconid, and positioned on the anterior basal part of the tooth lingual to the midline. /P4 is slightly shorter than /M1 on UCMP 134622 (Table 43), and the widest part of the tooth is at the junction of trigonid and talonid. The talonid is short but wide, with a low and small cusp situated at its posterior edge in the position normally occupied by a hypoconid.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In comparison to other "condylarths" found in the upper Hell Creek Formation, the premolars of Protungulatum gorgun are shorter and narrower than those of Ragnarok , but are longer and wider than those of Protungulatum donnae and Mimatuta morgoth . In basic morphology, the premolars of Protungulatum gorgun and P. donnae are similar, but the /P2-/P3 diastema is relatively longer in P. gorgun . Also, the /P4 of P. donnae is widest through the metaconid-protoconid (Archibald, 1982), while the /P4 of P. gorgun is widest more posteriorly at the talonid-trigonid contact. The /P4 talonid of P. gorgun is better developed (relatively wider and longer) and has a relatively larger hypoconid.
The lower molars of Protungulatum gorgun and P. donnae are similar morphologically, with a few exceptions. Lower molars of P. gorgun are larger, but similar in proportions (Tables 41, 44). The trigonid cusps of P. gorgun are relatively more bulbous and the paraconid and metaconid are more separated (Figures 24, 26). As in P. donnae , the paraconid of P. gorgun projects anteriorly, but does so to a greater degree. Unlike P. donnae , the paraconid in P. gorgun is usually shifted labially in relation to a line extended through the apices of the entoconid and metaconid. The hypoconid is larger relative to the other talonid cusps in P. gorgun (Figures 24, 26).
In comparison to Ragnarok , lower molars of Protungulatum gorgun are smaller and their trigonids are higher relative to the talonid. The trigonid and talonid cusps are less bulbous and the trigonid walls are much less expanded in P. gorgun . Also, the paraconid projects anteriorly in P. gorgun and does not (or projects very slightly) in Ragnarok .
The upper dentition of Protungulatum gorgun has never been described. Unfortunately, upper molars were not found in association with any of the mandibles referred to P. gorgun from McGuire Creek. Many isolated molars from McGuire Creek are similar to P. donnae , but are significantly larger and in the expected size range of P. gorgun . Many of these upper molars were collected from sites producing portions of
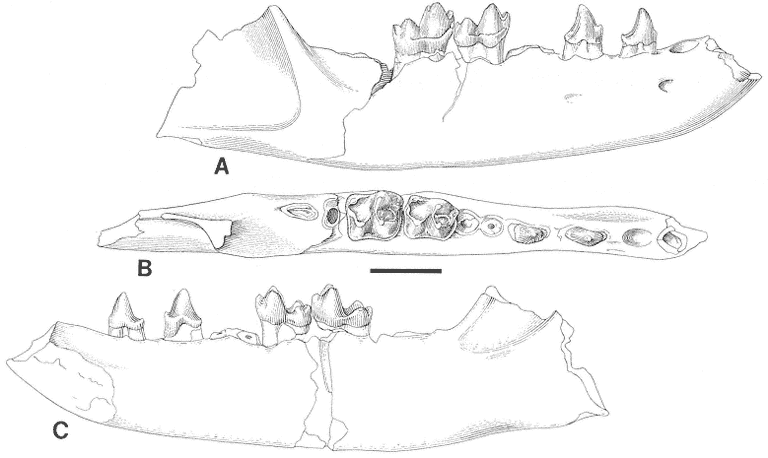
Figure 24
Protungulatum gorgun Van Valen. Right dentary with /P2-3, /M1-2, UCMP 134558,
locality V87037: (A) labial view, (B) occlusal view, (C) lingual view. Scale bar = 5 mm.
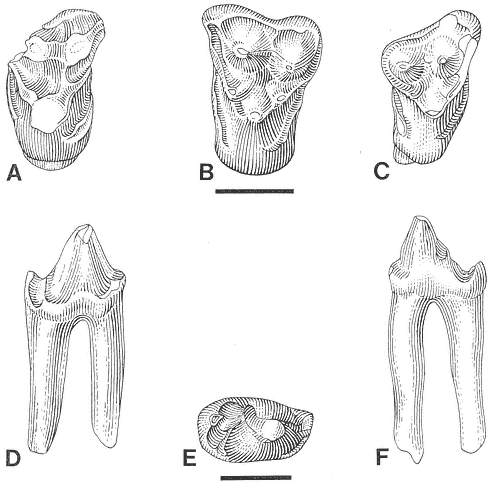
Figure 25
Protungulatum gorgun Van Valen. (A) Isolated left M1/, UCMP 132345, locality V87029,
occlusal view. (B) Isolated left M2/, UCMP 132507, locality V87035, occlusal view. (C)
Isolated right M3/, UCMP 133145, locality V87049, occlusal view. Scale bar = 4 mm.
Isolated right /P4 (associated with /M1-3 but molars not shown), UCMP 134622,
locality V87028: (D) labial view, (E) occlusal view, (F) lingual view. Scale bar = 3 mm.
lower dentitions that can be referred to P. gorgun with confidence. The only feature of the upper dentition mentioned in the diagnosis of P. gorgun concerns the protocone, "upper molars with rather massive protocone" (Van Valen, 1978, p. 52). The McGuire Creek fossils have fairly large protocones, but this character exhibits a wide range of variation. Therefore, reference of upper molars from McGuire Creek to P. gorgun must be regarded as tentative.
Both M1/'s from McGuire Creek referred to Protungulatum gorgun lack their parastylar region (Figure 25A). M1/'s of P. gorgun have a relatively larger protocone and a weaker postcingulum and hypocone than P. donnae . In other respects, they are basically a larger
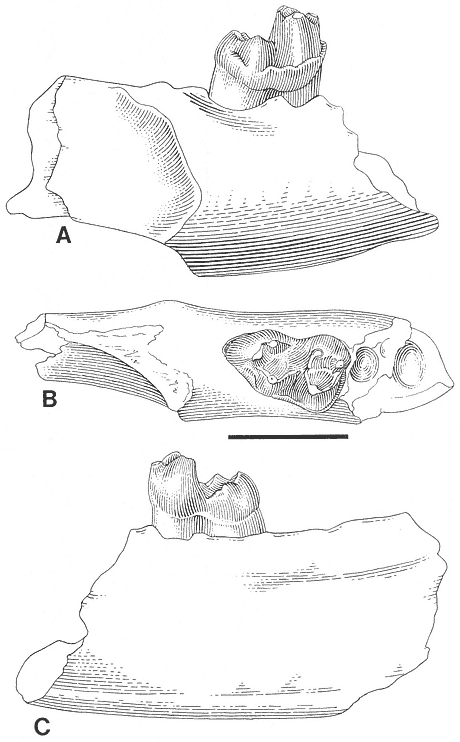
Figure 26
Protungulatum gorgun Van Valen. Right dentary fragment with /M3, UCMP 132318,
locality V87028: (A) labial view, (B) occlusal view, (C) lingual view. Scale bar = 5 mm.
version of M1/'s of P. donnae . The M2/ protocone of P. gorgun is high and large, which causes the labial and lingual protoconal slope to be steeper and longer than in M2/'s of P. donnae (Figure 25B). Also, the M2/ of P. gorgun is relatively wider. M3/'s of P. gorgun are larger than those of P. donnae , but otherwise are similar (Figure 25C). Dental measurements of P. gorgun are presented in Table 45. Upper premolars of P. gorgun were not identified.
Oxyprimus Van Valen, 1978
Oxyprimus erikseni Van Valen, 1978
Tables 46–48; Figure 27
Oxyprimus erikseni Van Valen, 1978, p. 53.
Type . UMVP 1561, isolated /M1 (Van Valen, 1978, pl. 1, fig. 5).
Type locality . Harbicht Hill, Hell Creek Formation, Montana.
Referred specimens . Dentary with /M3, UCMP 132453; 1 M2/ from loc. V86031. One M1/ from loc. V87028. Dentary with /M2-3, UCMP 132348; 1 M2/ from loc. V87029. One M1/ from loc. V87034. One /Mx from loc. V87035. One M1/ from loc. V87036. Dentary with /P2, /P4-/M2, UCMP 133068; 1 M1/ from loc. V87038. One /P4, 1 P4/ from loc. V87049. One M2/, 1 /M3 from loc. V87051. Dentary with /M1-3, UCMP 132350; 2 Mx/'s from loc. V87052. One M1/, 1 /M1, 2 /M2, 1 /M3, 2 M3/'s from loc. V87071. Two P4/'s, 1 DP4/, 2M1/'s, 1 M2/, 1 M3/, 2 /M1's, 1 /M2, 1 /M3, 2 /Mx's from loc. V87072. One M1/, 1 M3/, 1 /M1 from loc. V87074. One /M3 from loc. V87084. One /M2, 1 /M3 from loc. V87095. One P4/, 2 M1/'s, 2 /P4's, 1 /M3, 1 /Mx from loc. V87098. Three M1/'s, 2 /M3's from loc. V87151. One M1/, 2 Mx/'s, 1 /M3 from loc. V87153.
Localities . UCMP locs. V86031, V87028, V87029, V87034, V87035, V87036, V87038, V87049, V87051, V87052, V87071, V87072, V87074, V87084, V87095, V87098, V87151, and V87153.
Distribution . Upper Hell Creek and lower Tullock formations, Montana (both Puercan); possibly Ravenscrag Formation, Saskatchewan (Puercan).
Description . Reference of lower molars and premolars from McGuire Creek to
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Oxyprimus erikseni is based on comparison of /M1's to a cast of the type, UMVP 1561, an isolated /M1 from Harbicht Hill. Fortunately, two dentary fragments from McGuire Creek preserved /M1's with other parts of the dental arcade (Table 46; Figure 27C), and these specimens provide the basis for identification of isolated /P4's and /M2-3's. Dental measurements are given in Tables 46 and 47.
Upper and lower molars of Oxyprimus erikseni have not been found in association, and upper molars and premolars from Harbicht Hill, the type locality, have never been described. Isolated upper molars and premolars from McGuire Creek are referred to O. erikseni on the basis of comparison of these fossils with casts of O. galadrielae , a very similar species from Mantua Lentil. The upper dentitions of O. galadrielae and O. erikseni are difficult to distinguish (see discussion below), but upper molars and premolars referred to O. erikseni were collected from sites that produced portions of lower dentitions referred confidently to O. erikseni . The lower dentitions of O. galadrielae and O. erikseni are similar, but can be separated by a few characters (see discussion below). Therefore, although reference of these specimens from McGuire Creek to O. erikseni is probably correct, uncertainty exists. Dental measurements are given in Table 48.
The dentition of Oxyprimus erikseni was described by Archibald (1982), and his comments apply equally well to fossils referred to O. erikseni from McGuire Creek. A few modifications and additions are necessary. Archibald's (1982) identification of the tooth sites of three of the upper molars from Worm Coulee 1 (V74111) probably is incorrect. This conclusion is based on my comparison of these molars to specimens of Oxyprimus from McGuire Creek. UCMP 116513 and 116515 are actually M1/'s not M2/'s and UCMP 116511, as suspected by Archibald (1982), is a DP4/, not an M1/. Therefore, the M1/ and M2/ of O. erikseni require additional description. M1/'s and M2/'s of O. erikseni are similar to those of Protungulatum donnae (Archibald, 1982), but smaller. In contrast to P. donnae, Oxyprimus erikseni has a smaller parastylar lobe, a relatively larger hypocone, and the lingual slope of the protocone is steeper (Figure 27A-B). Also, weak internal conular wings are more commonly developed in O. erikseni than in Protungulatum donnae .
Discussion . A potential taxonomic problem exists in separating similar-sized species of Oxyprimus from Harbicht Hill and Mantua Lentil. The dentitions of O. erikseni from Harbicht Hill, Hell Creek Formation, Montana, and O. galadrielae and O. putorius from Mantua Lentil, Polecat Bench Formation, Wyoming, are similar.
Van Valen's (1978) diagnosis of each species is so brief that it became necessary to evaluate their validity. Identification of species is important, because regional biostratigraphic and biochronologic correlations are based in part on species-level differences within Oxyprimus . Van Valen (1978) only noted differences in metaconid shape as a means of separating O. galadrielae and O. erikseni . According to the diagnosis of each, O. erikseni has a conical molar metaconid and O. galadrielae has an elongate molar metaconid. Close scrutiny of the two species reveals that this is not the case. Molar metaconids of O. erikseni were found to be proportionately longer than or equal in diameter to those of O. galadrielae . Measurements were taken from the apex of the metaconid of /M1 to the transverse groove separating this cusp from the paraconid, and then were
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
compared with trigonid length. This ratio measures the elongation of the metaconid relative to length of the trigonid. Oxyprimus erikseni (n=3, including an epoxy cast of the type) has metaconid lengths of .78-.83 and elongation ratios of .44-.48. For O. galadrielae (from casts of PU 16712, PU 16863) these values are .71-.73 and .44-.45, respectively. On the basis of this small sample, the /M1 metaconid of O. erikseni appears to be equal to or slightly more elongate than metaconids of O. galadrielae . Inspection of /M2's and /M3's referred to O. erikseni from McGuire Creek and O. galadrielae from Mantua Lentil show the same trend. Therefore, molar metaconid elongation has questionable taxonomic value, and the diagnoses of O. galadrielae and O. erikseni are changed below to reflect this.
Because of the inadequacy of diagnoses of Oxyprimus galadrielae and O. erikseni , the possibility that these species may be synonymous was evaluated. Comparison of casts of
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
the lower dentition of O. galadrielae with specimens referred to O. erikseni from McGuire Creek and Harbicht Hill (including a cast of the type) revealed a few differences. O. erikseni has slightly narrower molar talonids than does O. galadrielae , both in proportion and in absolute size (Table 47). Also, in O. erikseni /P4's and /M1's are subequal in length, whereas in O. galadrielae /M1's are significantly longer than /P4's (Table 47). In most cases, O. erikseni has a larger /P4 metaconid than O. galadrielae although there is some degree of overlap. Otherwise the lower dentitions of these two species are similar.
Upper molars of Oxyprimus erikseni and O. galadrielae are also similar. Comparison of the type of O. galadrielae (PU 16866) to upper molars referred to O. erikseni from McGuire Creek reveal no significant differences. M2/'s from McGuire Creek appear to be slightly more transverse, but the difference is small, and proportionately they are identical (Table 48).
A second species from Mantua Lentil, Oxyprimus putorius , described briefly by Van Valen (1978), may be valid. Upper molars of O. putorius , known only from the type specimen (PU 16704), are less transverse than those of both O. galadrielae and O. erikseni
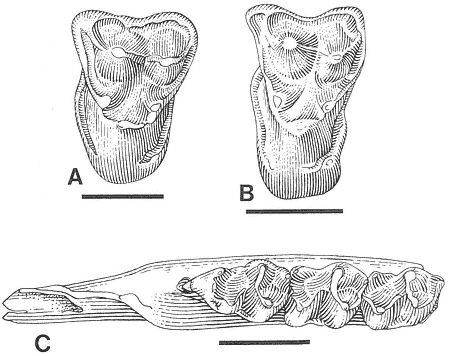
Figure 27
Oxyprimus erikseni Van Valen. (A) Isolated left M1/, UCMP 133447, locality V87072,
occlusal view. Scale bar = 2.5 mm. (B) Isolated left M2/, UCMP 134698, locality
V87072, occlusal view. Scale bar = 3 mm. (C) Right dentary fragment with /M1-3,
UCMP 132350, locality V87052, occlusal view. Scale bar = 3 mm.
(Van Valen, 1978), although proportionately there is little difference among upper molars of the three species (Table 48). The only known specimen of a lower dentition referred to O. putorius (PU 21099) is so poorly preserved that it cannot be used for comparative purposes. More material is needed to assess the validity of O. putorius .
In summary, O. galadrielae and O. erikseni are similar, but appear to be valid species that can be distinguished by differences in their lower dentitions. O. putorius is poorly known, and the validly of this species is questionable. A revised diagnosis of each species is given below:
Oxyprimus erikseni : /P4 metaconid usually large, narrower molar talonids than O. galadrielae , /P4 and /M1 subequal in length, upper molars more transverse than O. putorius .
Oxyprimus galadrielae : /P4 metaconid small, wider molar talonids than O. erikseni , /M1 longer than /P4, upper molars more transverse than O. putorius .
Oxyprimus putorius : upper molars less transverse than O. erikseni and O. galadrielae .
Subfamily LOXOLOPHINAE Van Valen, 1978
Ragnarok Van Valen, 1978
Ragnarok nordicum (Jepsen, 1930) Van Valen, 1978
Tables 49-52; Figures 28-30
Loxolophus nordicus Jepsen, 1930, p. 501-502.
Ragnarok harbichti Van Valen, 1978, p. 56.
Type . PU 13285, right maxilla with M1-3/ (Jepsen, 1930, pl. IV, fig. 1).
Type locality . Mantua Lentil, Polecat Bench Formation (Fort Union Formation), Wyoming.
Referred specimens . Dentary with /M2 talonid, /M3, UCMP 132594, from loc. V84151. Dentary with /M2-3, UCMP 132444; dentary with /P4-/M3, UCMP 132458; 1 M1/, 1 /M1, 2 /M2's, 2 /M3's from loc. V86031. Dentary with /M2-3, UCMP 132435; 1 /P4 from loc. V87033. Dentary with /M3, UCMP 132473; 1 /P4 from loc. V87034. Dentary with /M3, UCMP 132500, from loc. V87035. Two /M2's, 2 /M3's, 1 /Mx from loc. V87037. Associated /M1 and /M2 from loc. V87050. Dentary with /C, /P1-2, /P4, /M1-2, /M3 trigonid, UCMP 134797; maxilla fragment with P1/-M3/, UCMP 134694; maxilla fragment with M1-3/, UCMP 134693; 1 P4/ from loc. V87072. Dentary with /C, alveoli for /P1-3, UCMP 132804, from loc. V87088. Dentary with /M2 talonid, /M3, UCMP 132426, from loc. V87109. Dentary with /M2-3, UCMP 134592; dentary with /M2 talonid, /M3, UCMP 134591, from loc. V88041. One to three isolated teeth from 16 other localities.
Localities . UCMP locs. V84151, V86031, V87029, V87030, V87033, V87034, V87035, V87036, V87037, V87038, V87049, V87050, V87072, V87073, V87074, V87077, V87078, V87082, V87088, V87098, V87109, V87117, V87124, V87152, V87153, V88041, and V88044.
Distribution . Upper Hell Creek Formation, Montana, and Polecat Bench Formation (Fort Union Formation), Wyoming (both Puercan).
Discussion and Description . When Van Valen (1978) erected the genus Ragnarok , it included the genotypic species R. harbichti from Harbicht Hill, upper Hell Creek Formation, Montana; R. nordicum (Loxolophus nordicus , Jepsen, 1930) from Mantua Lentil, Polecat Bench Formation, Wyoming; and R. wovokae from Rock Bench, Wyoming. Later, Archibald (1982) named a new species, R. engdahli , based on isolated teeth from Worm Coulee 1 (V74111), lower Tullock Formation, Montana. Analysis of the type specimens and the hypodigm of these four species indicates that fossils from McGuire Creek probably represent a single population of Ragnarok that is similar to R. nordicum and R. harbichti . According to their diagnoses, Ragnarok nordicum and R. harbichti are similar and differ only in size of the molar talonids (see Van Valen, 1978). Specimens of R. harbichti from Harbicht Hill have slightly smaller /M1-2 talonids on average than the hypodigm of R. nordicum from Mantua Lentil, but proportionally (L/W-Tal) only the /M2 talonid of R. nordicum appears larger (Table 49). Because talonid width and total length of the /M2 appear to exhibit the greatest difference between the samples from the type localities of R. harbichti and R. nordicum , these values were subjected to T-tests. The differences in means of /M2 talonid width and /M2
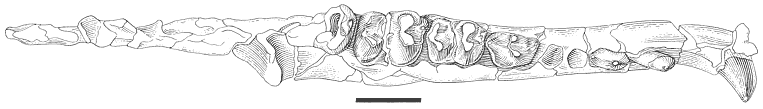
Figure 28
Ragnarok nordicum Jepsen. Right dentary with /C, /P1-2, /P4, /M1-2, partial trigonid of /M3, UCMP 134797, locality V87072, occlusal view.
Scale bar = 5 mm.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
length between the two samples were not significant (/M2 length: t=1.93, df=6, p=.10; /M2 talonid width: t=1.88, df=6, .20>p>.10). The Mantua Lentil and Harbicht Hill samples cannot be confidently separated by size of the molar talonids, and the diagnosis of each species is inadequate. A comparison of the remainder of the upper and lower dentitions of R. nordicum and R. harbichti reveals a lack of morphological characters that separate the species. On the basis of available information, R. nordicum and R. harbichti are considered synonymous, with R. nordicum having priority. Therefore, the sample of Ragnarok from McGuire Creek is referred to R. nordicum . It should be noted, however, that the sample sizes available for study from Harbicht Hill and Mantua Lentil are small (n=4 or less; Table 49).
Middleton (1983) has proposed that Ragnarok is synonymous with Baioconodon , and his views have gained acceptance (Sloan et al., 1986; Smit et al., 1987; Archibald and Lofgren, 1990). But until this synonymy is formally published, I will continue to recognize Ragnarok .
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
The dentition of Ragnarok nordicum has never been completely described. The only published morphological data are contained in a brief description of M1/-M3/ by Jepsen (1930) and Archibald's (1982) description of a possibly associated /M1-2 and a dentary fragment with damaged and heavily worn teeth (both from the lower Tullock Formation).
Description of the anterior lower dentition (/C, /P1-3) is based on a nearly complete but highly fractured dentary, UCMP 134797 (Figure 28). Two mental foramina are present, one below the posterior root of /P3 and another below the anterior root of /P2. The latter foramen is poorly preserved because of the highly fractured condition of the anterior part of the mandible. The canine is large, somewhat conical, with its apex extending much higher than the cusps of the premolars. The canine root is recurved beneath /P1. A single-rooted, slightly procumbent /P1 is separated from the canine by a small diastem. The /P1 has a small posterior basal cusp and a very small anterior basal cuspule, which is shifted slightly lingual to the midline of the tooth. The root of /P1 is the size of the posterior root of the double-rooted /P2. /P2 has a large central cusp which is higher than that of /P1, and a weakly developed talonid with a posterior basal cusp. An anterior basal cusp is not evident, but the enamel is slightly swollen in this region. The /P3 is broken off at root level and was not recovered; however, it had two roots. A diastem is not present between any of the premolars.
Description of remaining parts of the lower dentition is based on a much larger sample of associated (Table 50) and isolated teeth. The /P4 of Ragnarok nordicum has a massive protoconid and a large, low metaconid. The paraconid is large but is lower than the metaconid and is shifted lingually to the midline of the tooth (Figure 29). A protolophid is well developed on many specimens. The posterior labial cingulum is well developed, and a weaker anterior labial cingulum is present. These cingula are not connected across the protoconid. The talonid is well developed, with a large cusp in the position of the hypoconid (Figures 28, 29). The /P4 is widest at the trigonid-talonid junction.
The /P4 of Ragnarok is clearly larger than any of the other "condylarths" found in the upper Hell Creek Formation except Protungulatum gorgun . In comparison to P. gorgun , the /P4 of Ragnarok is longer and much wider, its protoconid is more bulbous, the posterior labial cingulum is better developed, and the paraconid and metaconid are relatively larger. Also, unlike Protungulatum gorgun, Ragnarok lacks premolar diastems.
Lower molars of Ragnarok nordicum have low trigonids with bulbous cusps that have greatly swollen lateral walls (see Figures 28, 29). The metaconid is slightly higher and larger than the protoconid. The paraconid is closely appressed to the metaconid and is much smaller than the other trigonid cusps. Also, the paraconid is shifted labially in relation to a line extending through the metaconid and entoconid. A small but distinct paralophid is present.
The talonid cusps are low but robust. The hypoconid is the largest cusp on /M1-2, with the entoconid and hypoconulid of subequal size. The hypoconulid is the largest cusp on /M3 and is usually inclined anteriorly. The hypoconid is slightly smaller, and the entoconid is anteroposteriorly elongated into a ridge. The labial cingulum is strong on all lower molars, and can be continuous from the paraconid posteriorly to the hypoconulid. The lingual cingulum is well developed at the talonid notch, but is rarely present elsewhere on the lingual side of the teeth. Distinct ectostylids are present on all lower molars, and the enamel in the hypoflexid region is commonly crenulated or rugose, especially on /M2.
In comparison to Protungulatum gorgun , the lower molars of Ragnarok are larger; have lower trigonids and more bulbous trigonid and talonid cusps; the trigonid walls of Ragnarok are greatly expanded; and the paraconid is closely appressed to the metaconid. The paraconid does not (or only slightly) projects anteriorly in Ragnarok .
Measurements of the lower dentition of R. nordicum from McGuire Creek are given in Tables 49 and 50.
A maxillary fragment with a nearly complete post-canine dentition (UCMP 134694, Figure 30B-D, Table 51) forms the basis for the description of the anterior premolars of Ragnarok nordicum . P1-P3/ were separated from the maxillary fragment during discovery. Unfortunately, the poor condition of the maxillary fragment is such that the anterior premolars cannot be remounted to ascertain their orientation or spacing.
The P1/ is single-rooted, but the crown is missing. Based on the size of the root, it was roughly comparable in size to the lower /P1. P2/ has two roots and a large anteroposteriorly elongated central cusp (Figure 30C). A prominent "metacrista" extends posteriorly to end in a small metastylar lobe. A small but distinct metastyle is present on a well developed labial cingulum. P3/ has a paracone and is more molariform than P2/, but still lacks any trace of a metacone (Figure 30D). The paracone is large, bulbous, and has a distinct metacrista. The metastylar lobe is large, with a prominent metastyle. The parastylar lobe is much smaller and has a small parastyle. The labial cingulum is continuous across the paracone. The protoconal lobe is missing, but the tooth narrows rather abruptly lingual to the paracone. If a protocone was present, it was small.
As with P3/, P4/ lacks a metacone but is more molariform because of the lingual extension of the protoconal lobe, which gives the tooth a transverse appearance (Figure
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
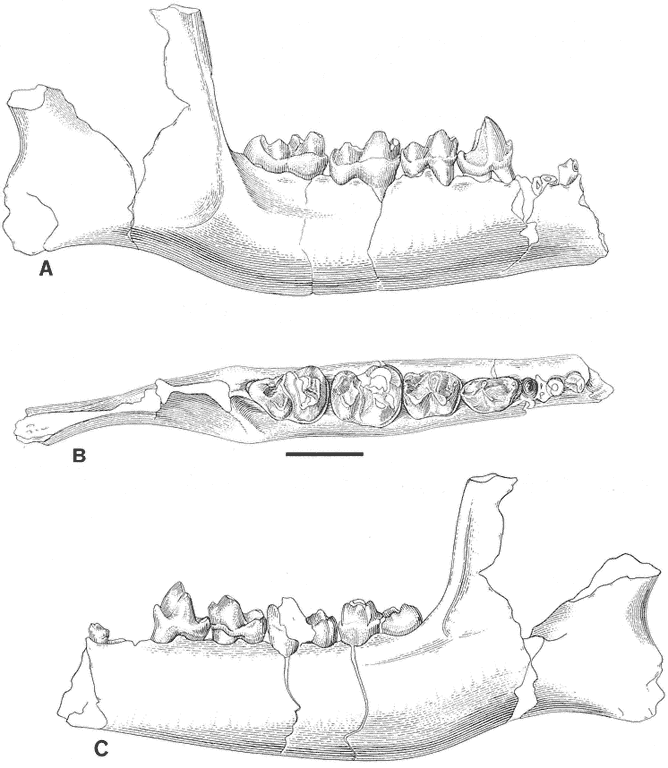
Figure 29
Ragnarok nordicum Jepsen. Right dentary with /P4, /M1-3, UCMP 132458, locality V86031:
(A) labial view, (B) occlusal view, (C) lingual view. Scale bar = 5 mm.
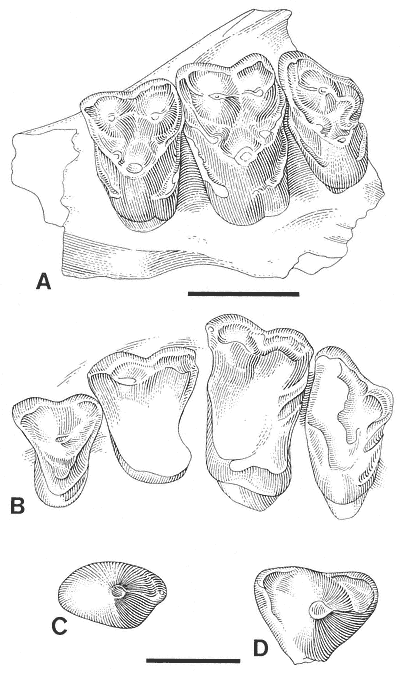
Figure 30
Ragnarok nordicum Jepsen. (A) Left maxillary fragment with M1-3/,
UCMP 134693, locality V87072, occlusal view. (B) Left maxillary fragment
with P2-3, M1-3, UCMP 134694, locality V87072, occlusal view of P4/,
M1-3/. Scale bar = 6 mm. Occlusal views of (C) P2/, and (D) P3/. Scale
bar = 3 mm.
30B). The protocone is large, and a weak pre- and postcingula are present. The paracone is larger than the protocone and has a metacristid extending posteriorly to a large metastylar lobe with a distinct metastyle. The parastylar lobe is smaller and has a small parastyle. The labial cingulum is continuous across the paracone, and an ectoflexus is well developed.
The upper molars of Ragnarok nordicum were described by Jepsen (1930), and only a few additional comments are necessary based on fossils from McGuire Creek. Upper molars have massive protocones and large conules lacking or with weak internal wings, except M3/, which usually has a strong premetaconule wing (Figure 30A). The M1/ has a weak paracrista and a distinct parastyle. A small mesostyle is usually present on M3/. Posterior lingual cingula are well developed, with distinct but small hypocones on M1-2/ (Figure 30A-B). M3/ lacks a hypocone. The lingual cingula are not continuous across the protocone in upper molars, with the exception of a single M2/.
In comparison to Protungulatum gorgun , the upper molars of Ragnarok nordicum have relatively more bulbous paracones and metacones. Also, their protocones are larger and much broader antero-posteriorly, giving the upper molars a more quadrate outline.
Measurements of associated and isolated upper molars and premolars of Ragnarok nordicum from McGuire Creek are given in Tables 51 and 52.
Family PERIPTYCHIDAE Cope, 1882c
Subfamily ANISONCHINAE Osborn and Earle, 1895
Mimatuta Van Valen, 1978
Mimatuta morgoth Van Valen, 1978
Tables 53-56; Figure 31
Mimatuta morgoth Van Valen, 1978, p. 62.
Type . UMVP 1560, right maxilla with M2/ (Van Valen, 1978, pl. 7, fig. 5).
Type locality . Harbicht Hill, Hell Creek Formation, Montana.
Referred specimens . Dentary with /P4, /M2-3, UCMP 132454, from loc. V86031. Dentary with /P4-/M2, UCMP 132340, from loc. V87029. One /P4, 1 M3/ from loc. V87037. Two /P4's from loc. V87038. One M3/ from loc. V87049. One /P4, 1 M2/ from loc. V87072. One M1/ from loc. V87077. Dentary with /P4-/M3, UCMP 134590, from loc. V87088. Two M1/'s, 2 M3/'s from loc. V87098. One M3/ from loc. V87101. Dentary with /C, /P1-4, /M1-3, UCMP 134589, from loc. V87123. Two /P4's, 1 M3/ from loc. V87151.
Localities . UCMP locs. V86031, V87029, V87037, V87038, V87049, V87072, V87077, V87088, V87098, V87101, V87123, and V87151.
Distribution . Upper Hell Creek Formation and lower Tullock formations, Montana (both Puercan).
Description . Only a few isolated upper molars from McGuire Creek are referable to Mimatuta morgoth . Archibald (1982) has adequately described the upper molars of M. morgoth , and only a few additional comments are required.
Archibald's (1982) admittedly tentative identification of the tooth site of UCMP 116518 is incorrect (this conclusion is based on comparison of UCMP 116518 to speci-
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
mens of Mimatuta morgoth from McGuire Creek). It is an M1/, not an M2/. Otherwise Archibald's description of upper molars of M. morgoth applies equally well to fossils from McGuire Creek.
The diagnosis of Mimatuta included the statement that the protocone was central in position (i.e. shifted labially) (Van Valen, 1978). Using a small sample (n=3), Archibald (1982) proposed that this relative labial shift of the protocone can be used to distinguish M. morgoth from the similar-sized species M. minuial and Protungulatum donnae . Recent work by Luo (1989, 1991), employing statistical analysis of the type of Mimatuta morgoth , an isolated M2/ from Harbicht Hill, and a large sample of upper molars of arctocyonids from Bug Creek Anthills, supports use of this character in separating Protungulatum donnae and Mimatuta morgoth . Analysis of the sample of M. morgoth from McGuire Creek (Table 53, n=9) also supports the use of this character in distinguishing species of "condylarths." The relative labial shift of the protocone is expressed in the A/W-A ratio (i.e., length of the lingual protoconal slope divided by the anterior width of the tooth), which is different in each of the three species. Examination of A/W-A ratios of fossils studied by Archibald (1982), the type of M. morgoth , and the McGuire Creek sample of the three species shows that M. minuial has higher and Protungulatum donnae lower A/W-A ratios than Mimatuta morgoth (Table 54). Therefore, this character appears to have diagnostic utility, as Archibald (1982) suggested.
Lower molars of Protungulatum donnae can be separated from those of Mimatuta by the labial shift of the paraconids in the latter (Archibald, 1982). But in contrast to the upper dentition, lower molars of M. morgoth are difficult to distinguish from those of M. minuial . The diagnosis of each species only recognizes differences in the size of the /P4 metaconid as the diagnostic character. The /P4 metaconid is "rather strong" in M.
| |||||||||||||||||||||||||||||||||||||||||||||
minuial and "rather weak" in M. morgoth (see Van Valen, 1978). Isolated and associated (with molars in dentaries) /P4's of Mimatuta from McGuire Creek can be separated into two distinct morphotypes, those with strong and those with weak metaconids. Five dentary fragments from McGuire Creek referable to Mimatuta , on the basis of the position of the molar paraconids, have the /P4 in place. Four of the five dentaries have weakly developed /P4 metaconids and are referred to M. morgoth (Table 55). The other dentary has a much more robust /P4 metaconid and is referred to M. minuial (Table 56). Comparison of molar morphology of dentaries referred to the respective species provides no clearly distinctive characters. Archibald (1982) suggested that the relative width of the molars that he tentatively referred to M. minuial (n=2) were greater than those of M. morgoth . Dentaries referred to each species from McGuire Creek on the basis of the development of the /P4 metaconid, show that molars of M. minuial and M. morgoth are approximately equal in relative width (Table 56) and cannot be distinguished on this basis. Casts of dentaries of M. minuial from Mantua Lentil, the type locality of the species, were measured and are included in Table 56. Comparison of molar talonid widths of M. minuial from Mantua Lentil and M. minuial and M. morgoth from McGuire Creek show that relative talonid width has questionable diagnostic value. Therefore, isolated lower molars of Mimatuta from McGuire Creek cannot be referred to either M. minuial or M. morgoth with confidence, and are assigned to Mimatuta species indeterminate.
Archibald (1982) has adequately described the lower dentition of M. morgoth except for the anterior parts of the mandible and its associated dentition. Based on more complete material from McGuire Creek, description of this part of the dentition is given below.
A well preserved dentary of M. morgoth with /C, /P1-4, /M1-3, (UCMP 134589,
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Figure 31, Table 55) shows that the canine has a vertically oriented, robust crown whose apex extends above /P1 and /P2 and equals the height of the central cusp of /P3. The canine has a weakly developed anterior ridge along the midline of the crown, and its root is strongly recurved posteriorly, extending beneath the root of /P1. The /P1 is slightly procumbent, and its central cusp is lower than that of /P2.
Figure 31
Mimatuta morgoth Van Valen. Left dentary with /C, /P1-4, /M1-3, UCMP 134589, locality V87123:
(A) labial view, (B) occlusal view, (C) lingual view. Scale bar = 4 mm.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Mimatuta minuial Van Valen, 1978
Tables 54, 56-57; Figure 32
Mimatuta minuial Van Valen, 1978, p. 62.
Type . PU 14211, left maxilla with P3-4/, M1-3/ (Van Valen, 1978, pl. 7, fig. 4).
Type locality . Mantua Lentil, Polecat Bench Formation (Fort Union Formation), Wyoming.
Referred specimens . One /P4 from loc. V87036. One /P4 from loc. V87038. Dentary with /P4-/M3, UCMP 134695; 1 /P4 from loc. V87072. Maxilla with M2/, UCMP 132308, from loc. V87091.
Localities . UCMP locs. V87036, V87038, V87072, and V87091.
Distribution . Upper Hell Creek and lower Tullock formations, Montana; Polecat Bench Formation (Fort Union Formation), Wyoming (all Puercan).
Description . The only specimen from McGuire Creek containing part of the upper dentition that confidently can be referred to Mimatuta minuial is a maxillary fragment containing a damaged M2/ (UCMP 132308). The M2/ is enamel-less (except for part of the trigon basin) and the parastylar lobe is missing. In spite of this damage, it is clear that the protocone of UCMP 132308 is shifted labially to a great degree, producing a high A/W-A ratio (Table 57), more so than in M. morgoth (Table 53). Also, remnants of the internal conular wings are preserved, and they appear to have been well developed, more so than in specimens of M. morgoth from McGuire Creek.
As discussed in the description of Mimatuta morgoth , the strong /P4 metaconid of M. minuial appears to provide the basis for separation of the two species. Casts of den-
Figure 32
Mimatuta minuial Van Valen. Right dentary fragment with /P4, /M1-3, UCMP 134695, locality
V87072: (A) labial view, (B) occlusal view, (C) lingual view. Scale bar = 4 mm.
| ||||||||||||||||||||
taries of the hypodigm from Mantua Lentil, the type locality of M. minuial , all have strong /P4 metaconids. The /P4 metaconid of UCMP 134695 from McGuire Creek is robust, and nearly equals the protoconid in height and size (Figure 32). Isolated /P4's from McGuire Creek have metaconids that are smaller and lower than their respective protoconids, but these are still much better developed than metaconids of M. morgoth .
As discussed in the description of M. morgoth , lower molars of M. minuial and M. morgoth apparently are not distinguishable unless they are associated with the /P4. Isolated lower molars and dentary fragments of Mimatuta without /P4's are referred to Mimatuta species indeterminate.
Mimatuta sp. indet.
Referred specimens . One /M2 from loc. V84151. One /M1, 1 /M3 trigonid from loc. V84193. Dentary with /M2 talonid, /M3, UCMP 132301, from loc. V84194. One /M1 from loc. V87030. One /M2 from loc. V87033. Dentary with /M1-2, /M3 trigonid, UCMP 132474, from loc. V87034. One /M1 from loc. V87035. Associated /P3-4, /M1-3, UCMP 134557; 1 /M2, 1 /M3 from loc. V87037. One /M1, 1 /M3 from loc. V87038. One /M2 from loc. V87049. One /M3 from loc. V87067. One /M2 from loc. V87071. One /M1, 1 /M3 from loc. V87072. One /M2 from loc. V87073. One /M1 from loc. V87074. One /M1, 1 /M2 from loc. V87077. Dentary with /M1-2, UCMP 132808; 1 /M2, 1 /M3 from loc. V87084. Dentary with /M1-2, UCMP 132317, from loc. V87095. One /M3 from loc. V87101. One /M1 from loc. V87151.
Localities . UCMP locs. V84151, V84193, V84194, V87030, V87033, V87034, V87035, V87037, V87038, V87049, V87067, V87071, V87072, V87073, V87074, V87077, V87084, V87095, V87101, and V87151.
Description . Includes dentary fragments and isolated lower molars that probably belong to either Mimatuta morgoth and M. minuial but are too fragmentary or incomplete to be referable to either species with confidence. The number of identifiable teeth of M. morgoth (upper molars, isolated /P4's, and dentaries with /P4's) from McGuire Creek greatly outnumber those referred to M. minuial . Therefore, it is likely that the majority of fossils placed in this category belong to M. morgoth .