Geographical Ecology of the Sacramento Valley Riparian Butterfly Fauna[1]
Arthur M. Shapiro[2]
Abstract.—The Sacramento Valley butterfly fauna is depauperate and strikingly uniform in the riparian corridor from Redding to the Sacramento-San Joaquin Delta. There are few obvious pre-American relicts and only one taxonomically recognized endemic, Battusphilenor hirsuta . Historical reasons for these conditions are discussed along with aspects of the biology of characteristic riparian species.
Introduction
The Sacramento Valley has the same reputation among Lepidopterists as among weekend recreation-seekers: a hot, dry, flat, uninteresting place between the coast and the Sierra Nevada that should be traversed as quickly and painlessly as possible. To some extent the Lepidopterological reputation, at least, is deserved.
Before 1968 no faunistic or ecological treatment of the Valley butterflies had appeared. In that year Opler and Langston (1968) published a faunistic analysis for Contra Costa County which included part of the Sacramento-San Joaquin Delta, and with it a very significant proportion of the Valley fauna. The entire butterfly fauna of the Valley consists of 65 species, of which only about 50 can be considered permanent residents under present-day conditions (Shapiro 1974, 1975). How rich or poor a fauna is this? It would seem rich to an Englishman; the British Isles, with 243,500 km2 (94,000 mi2 ) and infinitely more topographic and vegetational diversity, have about the same number of species as 19,400 km2 (7500 mi2 ) of Sacramento Valley (Ford 1975). On the other hand, by temperate North American standards this is clearly a poor fauna. A disturbed tidal marsh and adjacent waste ground covering some 104 km2 (40 mi2 ) near Philadelphia boast a fauna of 73 species (Shapiro 1970). Staten Island, New York, has 104 species in 180 km2 (70 mi2 ) (Shapiro and Shapiro 1973). Closer to home, Gates and Mix Canyons on the east slope of the Vaca Hills, Inner Coast Range, Solano County, California, have as many species as the entire Sacramento Valley (Shapiro unpublished). Almost any montane locality in northern California has more. The Trinity Alps and Mount Eddy have about 112 species (Shapiro etal . 1981); in the Sierra Nevada, Donner Pass (2,130 m. (7,000 ft.)) has over 100 species in about 31 km2 (12 mi2 ), giving it one of the richest butterfly faunas in the North Temperate Zone (Emmel and Emmel 1962; Shapiro unpublished).
Apart from species numbers, a fauna may also be examined by taxonomic composition. An "unbalanced" fauna is one in which one or a few taxonomic groups provide most of the species. When the Valley fauna is compared to others in California, it seems relatively balanced—there is a slight deficit of Lycaenidae, which tend to be specialists, and a corresponding surplus of Hesperiidae, which may be relative generalists (though their ecologies are mostly unstudied).
The reasons for local and regional differences in faunal richness and composition are partly historical and partly ecological.
In Miocene and Pliocene times much of the present Sacramento Valley was a large, shallow inland sea. As noted by Ornduff (1974), one consequence of the emergence of the Valley floor as a habitat for terrestrial plants was the evolution of new and distinctive taxa, mainly ephemeral annuals: "The surrounding upland areas had been occupied by plants for a much longer period of time than the Central Valley and support evolutionarily older plant species and plant communities". The evolution of distinctive plants and communities on the Valley floor was apparently not mirrored in its butterflies.
The pre-American Valley supported three principal community-types: bunchgrass/valley oak savanna, tule/cattail marsh, and riparian forest (Thompson 1961; Sculley 1973). Although the early explorers have left vivid accounts of northern California scenery and wildlife, our knowledge of the butterfly fauna of the pristine Valley is virtually non-existent. A great many
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Arthur M. Shapiro is Professor of Zoology and Lecturer in Entomology, University of California, Davis.
common California butterflies were described in Europe by J.B.A. de Boisduval, who received them from a Frenchman, Pierre J. Lorquin, who had come during the Gold Rush of 1849. Lorquin's Admiral, (Limenitislorquini ) a common riparian butterfly, is named after him. Nearly all we know of his travels is from the sketchy outline provided in de Boisduval's monograph (1869). He may have followed the Sacramento River north through the Valley to its headwaters (F. Martin Brown, inlitt .). De Boisduval says he "explored first all the environs of San Francisco, then the banks of the Sacramento and the Feather, . . . made trips into the Sierra Nevada range, even to the forests of the interior, braving the tooth of the bear and the fangs of the rattlesnake." Valley localities are mentioned in the text; for example Pierisprotodice is cited as "common enough in the Sacramento [district]".
Lorquin also furnished California butterflies to Felder in Austria and to a Philadelphia entomologist, Tryon Reakirt, who may have described a now-extinct riparian population of the Pierisnapi complex from near Stockton (see below). None of the early works gives a good picture of the Valley fauna as a fauna; almost nothing of an ecological nature was published on California butterflies before Tilden's (1959) landmark paper on Tioga Pass! We do know that no species was described from the Valley only to become extinct later, as happened in the San Francisco Bay Area. Whether this reflects a true absence of Valley endemics or only a failure to collect there is another matter. I imagine there was inadequate collecting—of a very limited fauna!
It seems apriori unlikely that the adaptive radiation of ephemeral annuals would have been paralleled in the butterflies. The timing of vernal annual life cycles is largely dependent on rainfall, which is notoriously unreliable and unpredictable. The normal evolutionary response of butterflies to these conditions is long-distance migration (Larsen 1976), as is seen in the Painted Lady (Vanessacardui ) in both the Old World and the New, and in a more disorganized way, in the lowland Orange Sulphur (Coliaseurytheme ), Checkered White (Pierisprotodice ), and others.
The surviving vernal-pool communities in the Sacramento Valley have their specialist bees, but no butterflies. The only grassland butterfly associated with a native annual is the Large Marble (Euchloeausonides ), which has a facultatively bivoltine race we infer was originally associated with the mustard, Thelypodiumlasiophyllum , but is now "domesticated" on weedy European Brassica . Even here, the Valley population—though phenotypically distinctive—has not been recognized taxonomically, and belongs to a Holarctic complex with vernal phenology in the Old World as well as the New.
The most striking, and rather surprising, absence in the Valley grassland fauna is that of a set of specialist Satyridae or Hesperiidae associated with the native bunchgrasses. The near-extermination of these grasses leaves little hope of finding relicts of a (totally hypothetical) pre-American fauna. Only the endemic California Ringlet (Coenonymphatullia california ), occurs today in disturbed grassland throughout the state, west of the Sierra-Cascade axis. Most of the Hesperiid fauna is weedy, associated with introduced grasses. At least one species, Lerodeaeufala , seems to be a recent introduction (since the 1940s). Hesperia juba , otherwise montane, occurs at low densities in the Bay Area and in the Delta.
Equally surprising is the lack of a distinctive fauna in the marshlands. They are older than the grasslands, basically the remnants of the old shallow sea, and better preserved. We have only the Yuma Skipper (Ochlodesyuma ), which feeds on a native strain of the common reed (Phragmitescommunis ), and is still numerous in the Delta. There are relict marsh skippers in the montane part of northern California, but they are rare and perhaps in decline. Overall, the marsh fauna in California is poor when compared to those in the East.
The riparian butterfly fauna is the best preserved in the Valley. Although we cannot be absolutely confident that nothing has been lost, the uniformity of the fauna throughout the region suggests that we are seeing most of it. Indeed, it might almost be said that the present Sacramento Valley butterfly fauna is a riparian fauna—albeit one that has expanded into irrigated agricultural and especially urban environments. The remainder of this paper surveys its derivation, distribution, composition, and prospects.
The Riparian Fauna Today
Sources
As California dried out over several million years, and the present Mediterranean climate became established, many plant species requiring summer moisture contracted their ranges to the banks of the major watercourses. This emerging riparian vegetation included woody taxa, ultimately derived from Axelrod's Arcto-Tertiary Geoflora, which otherwise could not have survived in the absence of summer rain (Raven and Axelrod 1978).
Precisely the same thing seems to have happened to the butterfly fauna. A sizeable proportion of the Valley fauna is of Arcto-Tertiary origin, with strong affinities to the Palearctic. This element is prominent in the riparian faunas. In most lowland California habitats butterfly emergences are concentrated in spring, coinciding with the simultaneous occurrence of good flight weather and lush vegetation. Most species are univoltine and have a summer dormancy, or diapause. This applies not only to Madro-Tertiary derivatives but Arcto-Tertiary ones as well, which may be multivoltine (e.g.,
Pierisnapi ) or early-summer univoltine (Satyrium spp., Glaucopsychelygdamus ) in more humid climates. Outside the riparian zone there are few multivoltine species, and those that occur are migratory or at least extremely dispersive.
Continuous breeding in summer requires continuous availability of host plants in suitable condition and adult food sources through the long rainless months. Comparing curves representing the number of species flying throughout the year at Valley and foothill locations, the difference in seasonal pattern is striking (fig. 1). It is due to the preponderance of multivoltine species in the riparian zone. These butterflies have no option of summer dormancy, though they do diapause overwinter. They have in effect been sheltered from the summer drought and have never evolved special physiological adaptations to cope with it. They are still doing what their forebearers did in a climate with summer rain.
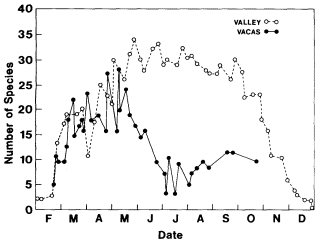
Figure l.
Number of species flying in the Vaca Hills and adjacent
Sacramento Valley during the 1972 season. The spring
peak in the hills is due to univoltine species, while most
Valley floor species are multivoltine. In the summer, annuals
are dried and dead. Perennials and woody plants have
ceased to add new growth. Thus both adult nectar and
larval food sources are unavailable in the Vaca Hills.
The preadapted character of this riparian fauna contrasts interestingly with the butterfly fauna of another region subjected historically to long-term drying: the Patagonian steppe. Patagonia has been deteriorating climatically since the Tertiary (Menendez 1972; Petriella 1972; Volkheimer 1971). The large rivers coming down from the Andes and crossing the vast treeless plateaus of Patagonia are fringed with a narrow band of riparian forest, but there is essentially no butterfly fauna there. Overall, the Patagonian butterfly fauna is far more depauperate and taxonomically unbalanced than that of the Sacramento Valley. The great majority of species are Satyrids that live not in the moist river bottoms but on the steppe proper, feeding on the bunchgrasses.
The historical basis for this apparent anomaly is straightforward. Given the past and present geography of the region, where would a riparian fauna be recruited from? Only the cool, humid Tertiary Nothofagus forests of the west slope of the Andes come to mind. But this is a very poor analogue of our Arcto-Tertiary sources. Arcto-Tertiary vegetation covered a vast area in the Northern Hemisphere and was extremely diverse. At that time the Nothofagus forests were already highly insular and much less diverse. Their extreme geographic isolation would have dictated an impoverished butterfly fauna even if the wet, cloudy climate had not. At the same time, an endemic butterfly fauna had differentiated in the treeless puna of the high Andes, and was able to colonize the cool-arid Patagonian steppe with little trouble. Even today, most of the shared taxa are undifferentiated even at the subspecies level. Clearly, the preponderance of riparian butterflies in the Sacramento Valley fauna does not represent a phenomenon inevitable in arid or semi-arid regions.
Although most of the Valley fauna seems to be of Arcto-Tertiary origin, two conspicuous species are not: Atlides halesus and Battusphilenorhirsuta . Atlides belongs to a purely Neotropical group. B . philenor has its closest relatives in tropical America but belongs to an old, pan-tropical group of Aristolochia -feeding Papilionidae best developed in southeastern Asia and the Australian region. Both B . philenor and its host plant, Aristolochiacalifornica may be relicts of Axelrod's Neotropical-Tertiary Geoflora, which is extinct at the level of woody plants. Alternately, they and A . halesus might have entered North America quite late in the "Great American Interchange" (Marshall 1981), as Anaea andria , Euptoietaclaudia , Eurema mexicana , and some other butterflies found in the eastern and southwestern United States almost certainly did. My own suspicion is that B . philenor , at least, is older.
Distribution
There are no endemic species in the butterfly fauna of the Sacramento Valley, and only one endemic subspecies (Battusphilenorhirsuta ). The riparian butterflies are largely widespread, generalized mesic species. Like the grassland and marsh faunas, they show little evidence of evolutionary activity. The fauna is extremely uniform from the head of the Valley to the Delta; the species lists for Turtle Bay (Redding) and Discovery Park (Sacramento) are nearly congruent. Nor is there north-south clinal variation or phenotypic differentiation, despite the increase in continentality in the climate of the north Valley.
These facts suggest genetic continuity in the riparian corridors, which have been fragmen-
ted only in recent historical time. Even in their present condition, these corridors are still probably adequate to permit the larger, more mobile species to maintain genetic continuity. Many riparian butterflies are excellent dispersers and are often seen as singletons many miles across hostile habitat from their nearest breeding colonies. B . philenor and Phyciodescampestris are good examples. Papiliorutulus has been documented as dispersing 8 km. (5 mi.).[3]
The question of whether these riparian corridors provided continuity across the Valley floor, conecting Coast Range and Sierran foothill populations, remains open. Most species are not phenotypically differentiated in the two foothill systems, and many of them occur in the isolated Sutter Buttes, even though there is no east-west continuity seen today (e.g., Papilio multicaudatus , Battusphilenor , Polygoniasatyrus , Vanessaatalanta ). Shapiro (1977) tells the strange story of Pieris napi castoria , described by Reakirt from Lorquin material possibly collected at or near the Valley town of Castoria (now French Camp, near Stockton); it is not known on the Valley floor today. The Chalcedon Checkerspot (Euphydryaschalcedona ) is a riparian/canyon species found in both sets of foothills which does not breed in the Valley today. It is phenotypically differentiated in both larval and adult characters between the two ranges, and has not been collected in the Sutter Buttes. Lorquin's Admiral shows weak phenotypic differentiation in northern California. Valley floor and Sierra populations are less orange below than those of the Coast Range, which resemble Trinity-Siskiyou area residents. Some of this variation may be under environmental control.
Some Distinctive Riparian Butterflies
Surveying the biology of some 50 taxa is beyond the scope of this paper. Readers desiring more complete documentation should consult Shapiro (1974, 1975). The following material presents information on the most distinctive components of the Sacramento Valley fauna.
Battus Philenor Hirsuta
The Hairy Pipevine Swallowtail, the Valley's only taxonomically recognized endemic entity, is also the most characteristic Valley and foothill riparian butterfly. It occurs everywhere that its sole host plant, Aristolochiacalifornica occurs. The species B . philenor occurs from Baja California and the Arizona desert to the mid-Atlantic coast. This subspecies, though weakly differentiated phenotypically, is unique in being completely disjunct from the rest of the species range. It is also unique in its population biology, sustaining extremely high densities in its larger colonies near Chico and in eastern Sacramento County. Its northern limit is near Dunsmuir, Siskiyou County. Except for an isolated record at North Bend, Oregon (Dornfeld 1980), it is completely restricted to the Sacramento Valley and adjacent canyons, and a few sites in the Bay Area.
The life-history of B . philenorhirsuta is of great ecological and evolutionary interest. Basically it is multiple-brooded, but part of the pupae in each generation go into diapause. Most of these do not hatch until the following spring, but a few emerge at irregular intervals the same season. The proportion diapausing, and the strength of the diapause, may be functions of the weather[4] rather than daylength, which determines these things in most mid-latitude butterflies.
The adaptive significance of this becomes apparent once the host relations of the butterfly are examined. Although large larvae can and do feed on mature Aristolochia foliage, young ones must begin feeding on the tender shoot tips, and it is here that the eggs are laid. In wet years the plants may put on some new growth all summer, and the butterfly can continue to breed. In very dry years breeding may become impossible as early as late June. The ratio of dormant to nondormant pupae in the very large spring brood thus represents a gamble on the condition of the plants a few weeks hence. The March–April flight is a mixture of adults from all the broods completed in the previous year. We are trying to learn how good this swallowtail is at predicting the condition of its host, and allocating its reproductive effort in different years.
Phyciodes Campestris
Not all Sacramento Valley populations of the Field Crescent occur in riparian forests; most are at the edges of such forests or of marshes. Its distribution largely coincides with that of its host plant, Asterchilensis . Aster is a prominent plant in mesic oldfield successions, but is rather rare and local in much of lowland California. Probably many colonies of both plant and butterfly remain undiscovered. In the southern Sacramento Valley (exclusive of the Delta, where it is relatively general), about six colonies are known. One of these (Willow Slough, north of Davis, Yolo County) has been under observation for 10 years. P . campestris is triple-brooded in the Valley and often very abundant where found. It overwinters as a larva, which can somehow survive being flooded for several weeks in winter.
Valley campestris show a distinctive summer phenotype not found elsewhere in the range, but we do not know if other populations would display it if reared under Valley conditions. The nominate subspecies with which our populations are lumped occurs from mesic sites in southern California (rare) north to Arctic Alaska. It also occurs east of the Sierra, as at
[3] L. Smith, personal communication.
[4] S.R. Sims, personal communication.
Mono Lake. Intervening between Great Basin and Sacramento Valley populations is a paler-colored entity, montana , endemic to montane and subalpine Sierran meadows. Typical campestris occurs in the same habitats in the Trinity Alps and Eddies. The complicated and often relictual distribution of this species in the far West seems to offer excellent opportunities for biogeographic interpretation, but its high vagility (i.e., potential colonizing ability) mandates caution.
Satyrium Californica
The California Hairstreak was not considered a riparian species until 1973, when the author found colonies in north Sacramento and eastern Yolo Counties, using valley oak (Quercus lobata ) as a host plant. No more such colonies have turned up, despite an assiduous search of oak groves (Walsh 1977). This may not mean no such colonies occur; Walsh's search was offseason, and the butterfly is easily missed—it spends much of its time in the trees, only coming down to feed around midday.
Although its existence was unsuspected before 1973, a valley oak ecotype of S . californica is hardly surprising. It commonly forms local ecotypes or ecological races, associated with various host plants of the genera Quercus and Ceanothus . At Rancho Cordova, Sacramento County, it is quite common—feeding on interior live oak (Quercus wislizenii ) and ignoring valley oak! In the subalpine zone on Packer's Peak in the Trinity Alps, it feeds on tobacco brush (Ceanothusvelutinus ). It is single-brooded everywhere, overwintering (and in the Valley, oversummering too!) as an egg.
Phenotypically, the valley oak ecotype is distinct from Coast Range foothill specimens, but not from the live oak-feeding Rancho Cordova population. It is not worthy of taxonomic recognition. So little valley oak vegetation remains that no one could be found to write a chapter on it for major synthesis of California vegetation studies.[5] Even if more than the three original colonies exist, the continuing deterioration of its habitat probably spells doom for this ecotype.
Lycaena xanthoides
The Great Copper presents the greatest mysteries in the Valley riparian fauna. Like Phyciodescampestris , it occurs in riparian broad-leaved vegetation, but also in moist grassland and at the edges of tule marsh. A glimpse at its northern California distribution (fig. 2) suggests that in the Valley it is restricted to the vicinity of the Sacramento River. There is no obvious reason for this. The butterfly is intensely colonial, and absent from a great many suitable-looking sites; yet it persists in the face of great disturbance. Its colonies occur on various soils, from sand to adobe clay; it is associated with at least six species of potential hosts (Rumex species, mostly introduced), none of which occurs in even a majority of its known colonies, except the very weedy R . crispus , whose range far exceeds the butterfly's.
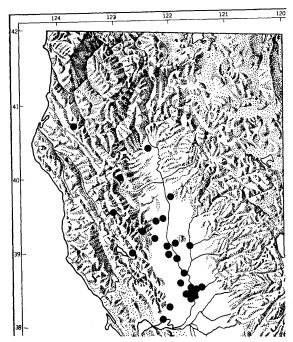
Figure 2.
Distribution of Lycaena xanthoides in and near the
Sacramento Valley, north of the Delta. Coastal and
Bay Area records, and the populations transitional
to L. editha from Dunsmuir to Weed, are omitted
(data in part from S.O. Mattoon; base map from
California Insect Survey).
Its distribution in the southern Valley has been carefully traced. It is common near Davis and Dixon (Solano County), and in the Yolo Bypass, West Sacramento, and Broderick (Yolo County). It crosses the Sacramento River at Discovery Park and ascends the American River to just beyond Highway 160, where it stops abruptly. As may be seen from the map, it has isolated (relict?) stations in interior Mendocino and Humboldt Counties, far from the Valley. It is common in the Delta and in vacant lots and marsh edges throughout the Bay Area, and extends to southern California. It is a montane species in the Tehachapis.
The Great Copper is completely unknown in the Sierra foothills and the adjacent east side of the Valley. In the montane and subalpine Sierra it is replaced by the smaller Lycaena editha , which extends northward at progressively lower elevations and appears to intergrade with L . xanthoides along Interstate 5 between Dunsmuir and Weed (Siskiyou County). L . editha is widespread in montane and subarid Ore-
[5] M.J. Barbour, Professor of Botany, University of California, Davis, personal communication.
gon, but true L . xanthoides occurs disjunctly in the Willamette Valley (Dornfeld 1980).
As with Phyciodescampestris , this species or species complex seems a good candidate for intensive biogeographic study. It has a single flight in late spring (May to early July). Univoltinism is characteristic of the entire subgenus, and winter is spent as an egg.
Prospects for the Future
Granted the continuing loss and fragmentation of riparian vegetation in the Sacramento Valley, the future of the butterfly fauna can be viewed in two lights. How likely is extinction at the level of populations or species? How are these extinction probabilities related to efforts to preserve riparian systems?
The generalized mesic character of the Valley riparian fauna argues against vulnerability on more than a local, or perhaps ultimately regional, scale. The fact that the fauna is largely derived from an Arcto-Tertiary biota adapted to mild climates with summer rain suggests that many of the species should still be common in such climates. In fact, we find them in the closest analogue available in California—the coastal fog belt. Polygoniasatyrus , for example, is very localized and rather rare in the Valley, but common and widespread in both Redwood and Douglas Fir associations coastwide. The Umber Skipper (Paratrytonemelane ) is an obligate riparian species in the Valley which can be found in every backyard in Berkeley.
Although several species like these might vanish from the Valley if riparian systems continue to disappear, they will leave large, healthy populations elsewhere. Many occur widely in the western half of North America, often with little geographic differentiation. Given the emphasis on endangered taxa (species and subspecies) in Federal conservation legislation, the Sacramento Valley riparian butterflies have little to offer conservationists looking for justifications for saving habitats. This is a pity, because butterflies have received a very favorable response in Washington. The sensitive entities are the colonial, endemic, or quasiendemic ones discussed in the preceding section. Two of these are not strictly riparian, and only one is a named subspecies.
Richard Arnold, in his Ph.D. dissertation,[6] studied the characteristics of most of the butterflies currently recognized as endangered or threatened. He found that low vagility, close association with a particular host plant and set of adult resources, and increasing fragmentation of the habitable range are attributes shared among them. However, most of the entities listed are Lycaenids, and a great many Lycaenids which are not presently endangered are local, sedentary, and monophagous. The critical variable is the amount (and contiguity) of breeding habitat. Many Lycaenids are potentially very vulnerable if their habitats become fragmented; other butterflies that resemble Lycaenids in population structure are also potentially in trouble. Let us see how these generalizations apply to the Valley riparian species.
Right now it seems unthinkable that Battus philenorhirsuta might become threatened; its annual population must run into the millions. Still, its reproductive capacity is low and it is completely dependent on a single host plant which is fairly habitat-specific. Notwithstanding these factors, a small colony (actually two colonies 2.4 km. (1.5 mi.) apart) has persisted on a handful of plants along the old channel of Putah Creek in Davis, which is now dry most of the year. These plants are growing in disturbed, mostly unshaded conditions and are subject to exceptional drought stress—yet they and their associated butterflies have remained essentially constant for the decade they have been under observation, even during the 1975–77 drought. In good years, surplus butterflies appear to emigrate from the colony. The Umber Skipper has disappeared from the same area since 1956. The persistence of this presumably relict population suggests that B . philenor can withstand much more fragmentation of habitat than it has yet had to face. However, as our most distinctive, spectacularly colorful, and taxonomically legitimate endemic it should be treated lovingly.
The other three species—Phyciodescampestris , Satyriumcalifornica , and Lycaena xanthoides —are colonial even without human interference. P . campestris fits all of Arnold's criteria for vulnerability except for its high vagility; individuals of both sexes are taken every year at Davis, 4.8 km. (3 mi.) from the nearest colony. They have indeed been taken at 1,525 m. (5000 ft.) on the Sierran west slope, within the range of P . c . montana and at least 80 km. (50 mi.) from the nearest known population in the Valley (Shapiro unpublished). The critical question here is how the spatial pattern of long-range dispersal maps onto the pattern of potentially colonizable sites.
Satyriumcalifornica and Lycaena xanthoides are quite sedentary, especially the former. Strays of xanthoides are recorded 16 km. (10 mi.) from breeding habitat (Shapiro unpublished). Colonies of xanthoides are numerous in the Davis-East Yolo-Sacramento area and gene flow remains likely. The status of the seemingly more isolated northern colonies is unknown. Its apparent rarity in the north Valley may merely reflect spotty collecting.
The long-term survival of the valley oak ecotype of S . californica is very unlikely. It perfectly matches Arnold's profile of an endangered butterfly. Had its phenotypic differentiation progressed to the same degree as the coastal dune endemics he studied, it would have been named and eligible for official Endangered
[6] Entomology, University of California, Berkeley, 1981.
status. But it did not. Even if additional colonies are located, the probabilities of inbreeding depression or random extinction are daunting. It requires oak reproduction for its own breeding. This imposes a limit on its tolerance of disturbance in the understory. It seems to be absent from those oak groves with the highest likelihood of preservation and selfrenewal. Its known sites are variously zoned for industrial development, and development as a waterfront regional park; and are located in a residential area where relict oaks have a low probability of surviving due to changes in drainage patterns.
The other side of the coin is the successful adaptation of a large part of the riparian butterfly fauna to the ersatz "riparian" systems in urban and suburban environments. Many of the most striking riparian butterflies, including Papilio rutulus , Nymphalis antiopa , Vanessaatalanta , and Atlideshalesus are breeding in street trees and gardens. I have observed 37 species of butterflies on my lot in one of the older subdivisions in Davis. None of the rarities breeds there, although I do get Phyciodescampestris in the garden regularly. So long as man provides mesic vegetation, the generalist multivoltine butterflies will use it.
The hope of finding relicts of the fauna of the pristine Valley has faded. We will probably never know what that fauna was like. Even so, there is plenty to be learned about the distribution and biology of the Valley riparian butterflies, and reason to hope that a few more Lepidopterists crossing the Valley enroute to somewhere else will be sufficiently intrigued to stop and take a look around.
Acknowledgments
I am indebted above all to Sterling O. Mattoon of Chico, who does collect in the Valley and who has graciously shared his data and perceptions with me. All of the data on figure 2 north of Knight's Landing and Marysville are his. I also thank Gary L. Rominger of the Sacramento County Department of Parks and Recreation for authorizing our ongoing studies of riparian butterflies along the American River Bikeway; everyone whose personal communications were cited in the text; and the Committee on Research and Department of Zoology, UCD, for funding work in the Sacramento Valley when they could have spent the money on research in Amazonia instead. Figure 1 was drawn by Ginny McDonald.
Literature Cited
Boisduval, J.B.A. de. 1869. Lépidoptères de la Californie. Annales de la Société Entomologique de Belgique 12:5–95.
Dornfeld, E. 1980. The butterflies of Oregon. 276 p. Timber Press, Forest Grove, Oregon.
Emmel, T.C., and J.F. Emmel. 1962. Ecological studies of Rhopalocera in a High Sierran community—Donner Pass, California. I: Butterfly associations and distributional factors. Journal of the Lepidopterists' Society 16(1):23–44.
Ford, E.B. 1975. Butterflies. Revised edition. 368 p. Collins-Fontana New Naturalist Series, London, England.
Larsen, T. 1976. The importance of migration to the butterfly faunas of Lebanon, East Jordan, and Egypt. Notulae Entomologicae 56:73–83.
Marshall, L.G. 1981. The Great American Interchange—an invasion-induced crisis for South American mammals. p. 133–230. In : M.H. Nitecki (ed.). Biotic crises in ecological and evolutionary time. 301 p. Academic Press, New York, N.Y.
Menendez, C.A. 1972. Paleofloras de la Patagonia. p. 129–184. In : M.J. Dimitri (ed.). La región de los Bosques AndinoPatagónicos. INTA, Buenos Aires, Argentina.
Opler, P.A., and R.L. Langston, 1968. A distributional analysis of the butterflies of Contra Costa County, California. Journal of the Lepidopterists' Society 22(2):89–107.
Ornduff, R. 1974. Introduction to California plant life. 152 p. University of California Press, Berkeley.
Petriella, B. 1972. Estudio de maderas petrificadas del Terciario Inferior del area de Chubut Central. Revista del Museo de La Plata, nueve serie, Paleontología 6:159–254.
Raven, P.H., and D.I. Axelrod. 1978. Origin and relationships of the California flora. 134 p. University of California Publications in Botany, vol. 72.
Sculley, R. 1973. The natural state. Davis New Review 1(1):3–13.
Shapiro, A.M. 1970. The butterflies of the Tinicum region. p. 95–104. In : Two studies of Tinicum Marsh. 124 p. Conservation Foundation, Washington, D.C.
Shapiro, A.M. 1974. The butterfly fauna of the Sacramento Valley, California. Journal of Research on the Lepidoptera 13:73–82, 115–122, 137–148.
Shapiro, A.M. 1975. Supplementary records of butterflies in the Sacramento Valley and Suisun Marsh, lowland central California. Journal of Research on the Lepidoptera 14: 100–102.
Shapiro, A.M. 1977. Photoperiod and temperature in phenotype determination of Pacific Slope Pierini: biosystematic implications. Journal of Research on the Lepidoptera 16:193–200.
Sharpiro, A.M., and A.R. Shapiro. 1973. The ecological associations of the butterflies of Staten Island (Richmond County, New York). Journal of Research on the Lepidoptera 12:65–128.
Shapiro, A.M., C.A. Palm, and K.L. Wcislo. 1981. The ecology and biogeography of the butterflies of the Trinity Alps and Mount Eddy, northern California. Journal of Research on the Lepidoptera 18:69–152.
Thompson, K. 1961. Riparian forests of the Sacramento Valley, California. Annals of the Association of American Geographers 51:294–314.
Tilden, J.W. 1959. The butterfly associations of Tioga Pass. Wasmann Journal of Biology 17:249–271.
Volkheimer, W. 1971. Aspectos paleoclimatológicos del Terciario Argentino. Revista del Museo de Ciencias Naturales "Bernardino Rivadavia," Buenos Aires, serie Paleontológica 1:243–262.
Walsh, J.B. 1977. Report on the distribution and status of Quercuslobata groves in California which may support an ecotype of Satyrium californica . Atala 5:7–12.