Ringtail Distribution and Abundance in the Central Valley of California[1]
Linda Belluomini and Gene R. Trapp[2]
Abstract.—The presence of ringtails (Bassariscusastutusraptor ) in riparian systems of the northern Central Valley is documented in this study, constituting a range extension. Ringtail densities 2.5 to 5 times greater than reported in the literature for other habitats were recorded, viz . 10.5 to 20.5 per 100 ha. (26.7 to 52.8 per mi2 ). Habitat composition was examined and related to the densities obtained. A direct relationship is suggested between ringtail density and community productivity and structural complexity.
Introduction
Ringtails (Bassariscusastutus ) are distributed throughout 11 southwestern states and portions of Mexico (Long and House 1962; Hall 1981). Throughout their range they are typically associated with boulder-strewn chaparral, chaparral interspersed with evergreen woodland, oak woodland, and scrub vegetation of various types (Seton 1929; Grinnell etal . 1937; Taylor 1954; Davis 1960; Hall 1981). Within California, ringtails inhabit the "Upper Sonoran Life Zone" on Pacific drainage slopes from the Oregon border to Mexico and the mountain ranges east of the Southern Sierra Nevada (Grinnell etal . 1937). Although acknowledging the presence of ringtails (the race B . a . raptor ) in streamside vegetation to about the 160-m. (500ft.) elevation, Grinnell etal . (ibid .) apparently found no evidence to suggest that ringtails occurred in this vegetation-type on the floor of the Central Valley. However, they did report ringtail observations for the Sacramento River in southern Tehama County, in the northern end of the Valley. Their belief that ringtails were restricted to the surrounding foothills and mountains is apparent in their distribution map.
The first published report of ringtails occurring on the Valley floor was made by Naylor and Wilson (1956). They observed ringtails in Wood Duck (Aixsponsa ) nest boxes in Butte Sink, along Butte Creek, 4.8 km. (3 mi.) north-east of Colusa, Colusa County, on the floor of the Sacramento Valley. Notwithstanding the recognized occurrence of ringtails in Butte Sink (Hall and Kelson 1959; Schempf and White 1977; Hall 1981), reviews of ringtail distribution (op. cit.; Ingles 1965) subsequent to Grinnell etal . (1937) did not modify the range of ringtails in California relative to the Central Valley.
In December 1971, Dr. Dallas Sutton[3] informed Trapp about the ease of collecting ringtails along the Sacramento River southwest of Chico for his mammal collection at California State University at Chico. Subsequently, two recent mammal inventories suggested that ringtails may occur in riparian vegetation associated with the network of drainages in the Central Valley. Stone (1976) detected an abundance of ringtails at certain sites along the Sacramento River in Tehama County at localities approximating those reported to have ringtails by Grinnell etal . (1937). Also, while conducting a mammal census of the Bobelaine Audubon Sanctuary on the Feather River at its confluence with the Bear River in Sutter County, during the spring of 1978, members of the Ecological Research Society of California State University, Sacramento, livetrapped eight ringtails. These data also suggested ringtail densities in riparian vegetation greater than those reported in the literature for other types of systems (ibid .; Taylor 1954; Trapp 1978).
The present study had three objectives: 1) to refine earlier distribution studies of the ringtail in California with respect to the Central Valley; 2) to verify preliminary observa-
[1] Paper presented at the California Riparian Systems Conference. [University of California, Davis, September 17–19, 1981].
[2] Linda Belluomini is Resource Specialist, Natural Resources, Marine Corps Base, Camp Pendleton, Calif. Gene R. Trapp is Associate Professor of Biological Sciences, California State University, Sacramento.
[3] Dr. Dallas Sutton, Emeritus Professor, California State University, Chico. Personal conversation.
tions which indicated desities much higher in riparian vegetation of the Central Valley than reported in the literature for other vegetation-types occupied by ringtails; and 3) to examine the composition of the riparian plant community associated with sites occupied by ringtails.
Methods and Materials
Ringtail distribution in the Central Valley was evaluated using five methods: 1) a literature review was conducted; 2) a furbearer observation questionnaire was circulated statewide; 3) locality data were gleaned from museum specimens; 4) knowledgeable persons were contacted; and 5) ringtails were captured by livetrapping.
Furbearer observation reports requesting specific information on ringtail sightings were distributed to appropriate Federal, State, and local agencies (e.g., wildlife refuges, sanctuaries, and preserves; California Department of Fish and Game biologists and wardens), as well as licensed trappers throughout California. Letters requesting pertinent locality information were sent to selected college and university museums which might have on deposit ringtail skins and/or skulls collected in the Valley. Personal contact was initiated with persons known to be familiar with wildlife of the Central Valley. Livetrapping was conducted during the summer and fall of 1978 and 1979 at nine sites in the Sacramento Valley to determine if ringtails were present.
Ringtails were easily captured using singledoored 9x9x26-in. and double-doored 6x6x24-in. galvanized, wire mesh livetraps.[4] Several baits (strawberry jam, raisins, muskrat meat) and ringtail lure were used to attract ringtails into the traps. Strawberry jam was the most effective attractant.
Data used for computation of density were collected by livetrapping, tagging, and recapture at five of the nine sites mentioned above. Livetraps were placed in a nonrandom manner (i.e., adjacent to ringtail scats; on, or alongside logs, etc.); throughout a portion of a particular study site. When recaptures indicated that all individuals within the immediate vicinity had been caught, the traps were moved variable distances away from that location until unmarked individuals were captured. This direct count process was repeated until presumably all ringtails within a study site were trapped. Aerial photographs were planimetered to determine the area of each study site, excluding habitats where ringtails were not captured or areas thought to be non-ringtail habitat. The number of ringtails per unit area was determined by dividing the total number captured at a study site by the total area, giving what amounts to an "ecological" density (Odum 1971; Smith 1980).
To facilitate handling of ringtails, Tranvet 25[5] was used during the early stages of this study. The Tranvet was orally administered using an eyedropper. Dosages ranging from 0.16 ml. to 0.30 ml. were given to individuals weighing from 870 gm. to 1300 gm.
The effect of the Tranvet was not consistent. Hence, an alternative and more effective means of subduing the ringtails was sought. Ketamine hydrochloride,[6] a derivative of phencyclidine, was successfully used. Dosages approximating 0.01 ml. per 100 gm. of body weight were sufficient to sedate ringtails in 1-4 minutes for the 10–20 minutes necessary to tag, measure, weigh, and examine them. Recovery from the drug occurred 30–60 minutes after injection.
Each captured ringtail was weighed (to the nearest 10 gm.) in a burlap sack. Data on the following variables were taken: 1) sex; 2) lengths of tail, body, right hind foot, right ear-to-notch, and right ear-to-crown; 3) circumferences of neck and cranium (all measurements recorded in mm.); 4) pelage condition; and 5) body temperature. A numbered ear tag[7] was fixed to the medial proximal edge of the right ear. Animals were released at the point of capture, after recovery from the drug.
The vegetation on one of the study sites (Site V) was analyzed by the plotless pointquarter and quadrat methods (Cottan and Curtis 1956; Cox 1976). Motroni (1978, 1979) used these techniques to collect data on the composition of riparian vegetation at four of the study sites (Site I, II, III, IV). Overstory, midstory, and understory layers were distinguished (as described by Motroni 1978) and analyzed separately. The plotless point-quarter method was used to collect data on overstory and midstory vegetation. The quadrat method was used to evaluate the understory. In this study, the boundaries of the Central Valley correspond to those of "California Prairie" described by Küchler (1977), excluding that found within Monterey County.
Study Sites
Ringtail densities were determined at five study sites in riparian vegetation of the Sacramento Valley (fig. 1).
Study Site I
Preliminary trapping efforts were conducted at the Bobelaine Audubon Sanctuary, located on the Feather River near its confluence with the Bear River, approximately 3.2 km. (2.0 mi.) north of the town of Nicolaus, Sutter County, Califor-
[4] Tomahawk Live Trap Co., P.O. Box 323, Tomahawk, Wisconsin 54487.
[5] Propio-promazine hydrochloride, 100 mg./ml., Diamond Laboratories, Inc., Des Moines, Iowa.
[6] Vetelar, Parke-Davis & Co., Detroit, Michigan 48232.
[7] No. 1, monel metal, Jiffy wing bands from National Band and Tag Co., Newport, Kentucky.
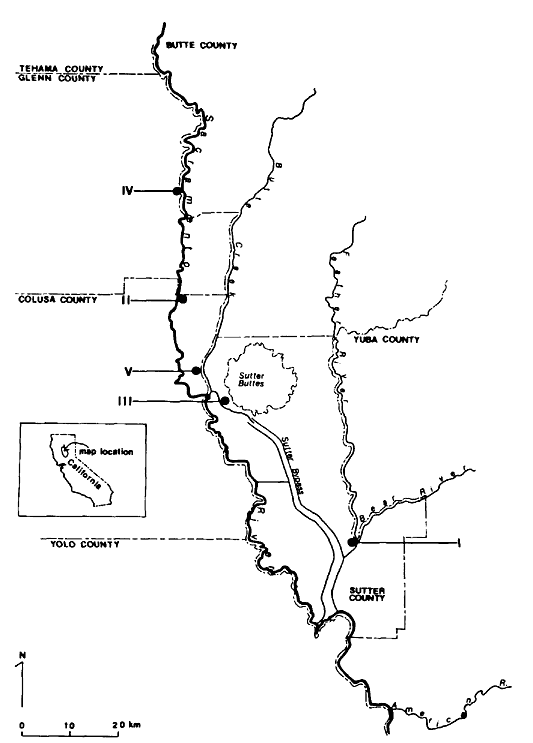
Figure l.
Ringtail study site locations in the Sacramento Valley, California.
nia (38° 56' 15" north, 121° 35' 45" west). The sanctuary is bordered on the east by the Feather River, on the west by walnut orchards, and on the north and south by riparian woodland. A slough traverses the western edge of the sanctuary and at one point widens into a 4.7 ha. (11.5 ac.) pond.
An extensive vegetation survey of the study site was conducted by Motroni (1979). Five major plant associations were recognized: 1) river grassland; 2) riparian shrub; 3) oak woodland; 4) cottonwood forest; and 5) riparian forest. It was within the last two plant associations and primarily in the riparian forest bordering the slough and pond that the majority of ringtails was livetrapped. These areas comprise approximately 69 ha. (172 ac.). Motroni (ibid .) found the overstory of the riparian forest to be dominated by cottonwood (Populusfremontii ). Other species present included box elder (Acer negundo ), valley oak (Quercuslobata ), and black walnut (Juglanshindsii ). The dense midstory was dominated by wild grape (Vitiscalifornica ), poison oak (Rhusdiversiloba ), Mexican tea (Chenopodium ambrosioides ). Other species present included blackberry (Rubusvitifolius ), coyote bush (Baccharispilularis var. consanguinea ), and wild rose (Rosacalifornica ). The understory was dominated by bermuda grass (Cynodondactylon ).
Study Site II
This 88-ha. (218-ac.) site is located on the Henry Womble property on the east bank of the Sacramento River 2.4 km. (1.5 mi.) south of Princeton, Colusa County, California (39° 22' 30" north, 122° 00' west). This is virtually a riparian island bordered by the Sacramento River and surrounded on three sides by agricultural fields. There are two small ponds, each less than 0.8 ha. (2 ac.), on the north side of the study site. An oxbow borders the east edge and curves west through the site where it eventually meets the Sacramento River. The vegetation has been described by Motroni (1978). Cottonwood was the dominant overstory tree, with willow (Salix sp.), valley oak, black walnut, and Oregon ash (Fraxinuslatifolia ) also present. Wild grape, blue elderberry (Sambucuscaerulea ), black walnut, and box elder formed a dense, often impenetrable midstory. Wild grape dominated the understory much the same as it did the midstory. Also abundant in this layer were blackberry and poison oak.
Study Site III
This 19-ha. (48-ac.) site, the Butte Slough State Wildlife Area, is located in Sutter County 26.4 km. (17 mi.) west of Yuba City (39° 9' north, 121° 53' west). Butte Slough forms the western boundary; agricultural land borders the site to the north and south, and a narrow band of oak woodland savannah was found along the east edge (beyond which is agricultural land). A 1.2-ha. (3-ac.) pond is located near the east edge of the study site.
This site differs markedly in vegetative composition and physiognomy from all other study sites. The riparian forest was a non-contiguous stand. Motroni (1978) found the overstory dominated by cottonwood with some willow and traces of valley oak. The relatively open midstory was dominated by Oregon ash. Buttonbush (Cephalanthusoccidentalis var. californicus ), valley oak, and box elder were also present in this layer. Poison oak, wild grape, and cocklebur (Xanthium strumarium var. canadense ) dominated the understory.
Study Site IV
This 76-ha. (187-ac.) site is on the west bank of the Sacramento River 6.9 km. (4.3 mi.) north of Glenn County, California (39° 35' north, 122º 00' west). It was formerly owned by Louis Heinrich, but now is a state wildlife area, and is bordered by the Sacramento River to the east and by agricultural land on three sides. A slough branches off of the river and borders the west side of the study area. Cottonwood and willow dominated the overstory, with box elder, black walnut, and sycamore (Platanus racemosa ) also present in this layer (ibid .).
Blue elderberry, willow, box elder, and black walnut combined to form a dense midstory, much like that found at Study Site II. Mugwort (Artemesiadouglasiana ), box elder seedings, wild cucumber (Marahfabaceus ), bed straw (Galiumaparine ), poison oak, black walnut seedlings, various grasses, wild grape, and blackberry formed the understory. Although not included in the vegetation survey conducted by Motroni (ibid .), approximately one-fourth of the study area was composed of a sparse stand of cottonwood with wild grape forming lianas similar to that described by early explorers of the Sacramento Valley.
Study Site V
This 82-ha. (202-ac.) site is 4.8 km. (3.0 mi.) northeast of Colusa, Colusa County, California (39° 16' north, 121° 57' west). It is adjacent to Butte Lodge Outing Duck Club and adjoins 0.2 km. of Butte Creek. A narrow slough runs the length of this riparian forest. Willow and valley oak dominated the overstory, with cottonwood, Oregon ash, and box elder contributing to form a dense canopy. Density of the midstory, dominated by poison oak, was variable. Oregon ash, box elder, buttonbush, and valley oak were also relatively abundant in this layer. Areas immediately adjacent to water were occupied by extensive stands of wild grape and willow forming a dense, nearly impenetrable midstory similar to Study Sites I, II, and IV. The closed canopy precluded much development of an understory. Poison oak was the dominant understory species, with blackberry, wild grape, mugwort, buttonbush, valley oak seedlings, and various grasses also present.
Results and Discussion
Ringtails were livetrapped or reported to exist at numerous locations in the Central Valley, including five of the nine sites sampled during this study (table 1; fig. 2, 3). Sightings ranged from the northernmost portion of the Valley near Red Bluff, Tehama County, to Stockton, San Joaquin County. These data represent a range extension, notwithstanding Naylor and Wilson (1956).
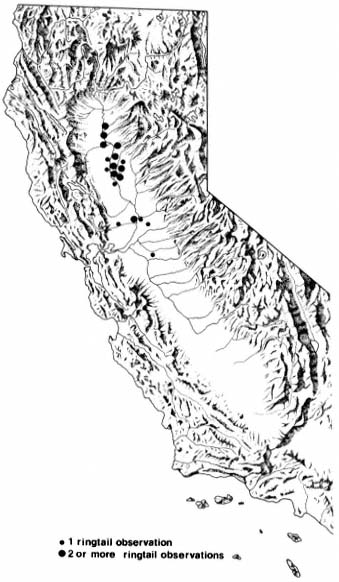
Figure 2.
Sites at which observations of ringtails have been
made within the Central Valley of California,
showing spatial relation to the rest of the state.
With few exceptions, ringtails were found to be associated with remnant stands of riparian forests bordering waterways such as the American River, Sacramento River, Feather River, Butte Creek, and Butte Slough. No ringtails were captured or reported from open, park-like stands of valley oak woodland.
Ringtails were also captured by Trapp in riparian vegetation in the Sutter Buttes, a small, isolated mountain range in the lower central portion of the Scramento Valley. Walt Anderson[8] reported that ringtails or their signs have been seen in other vegetation-types in the Sutter Buttes, including the blue oak woodland.
It is unlikely that ringtails have dispersed into the Valley since the statewide survey by Grinnell etal . (1937). In fact, evidence, apparently overlooked by Grinnell etal ., exists in the Museum of Vertebrate Zoology, University of California, Berkeley, from the 1930's which indicates ringtails were present at the Sutter Buttes (table 1, Sutter County). In their discussion of ringtail habitat preference, Grinnell etal . (ibid .) indicated that outside of the preferred "brushy or chaparral type" habitat, ringtails may also be ". . . found along streams (where) their range extends down to 500
[8] Walt Anderson. 1981. Biologist, Colusa, California. Personal communication.
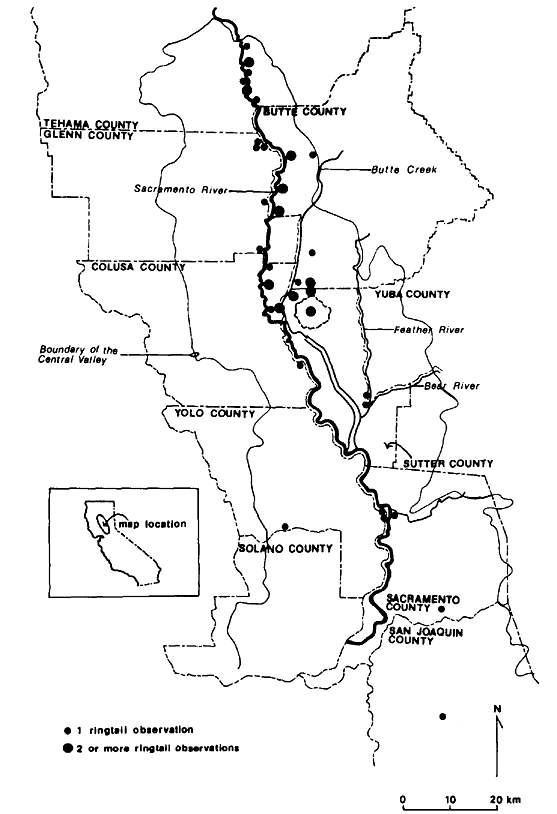
Figure 3.
Observation sites of ringtails in the Central
Valley of California, showing relation to
county boundaries and other local features.
ft. or to the mouths of canyons". Careful examination of the Grinnell range map (ibid .) revealed five ringtail sightings extending from near Red Bluff to the southern border of Tehama County. These sightings, which were just below 160 m. (500 ft.) and within the limit of California Prairie (Küchler 1977, i.e. the Central Valley), appeared to follow the Sacramento River. It is not clear why ringtails were not expected to occur in similar habitat below 160 m. It appears that ringtails were largely overlooked within the Central Valley and that there was simply a lack of effort by other investigators to document their presence prior to the current study.
The lack of sightings for the southern half of the Central Valley could be related to several factors. Historically, there was considerably less riparian vegetation here than in the northern Central Valley (ibid .; Roberts etal . 1977). A large portion of the remaining riparian vegetation has been eliminated in the path of agricultural expansion. Limited habitat availability, coupled with a less intensive survey effort by us in that area may explain why the range appears restricted to the northern portion of the Central Valley.
Trapping efforts undertaken to determine the abundance of ringtails in riparian forests of the Sacramento Valley produced reasonably consistent results where habitat features and trapping efforts were similar (table 2, 3). At study sites II, IV, and V densities of ringtails were 20.5, 17.1, and 19.5 individuals per 100 ha., respectively. Study sites I and II, where habitat features were different or where trapping effort was significantly less, exhibited densities of 11.4 and 10.5 ringtails per 100 ha.
At study site III, trap nights per ha. and trap nights per capture exceeded that of other sites (table 3); however, ringtail density at this site was lowest. There are probably a
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
variety of environmental factors involved in this lower value. This site, which borders Butte Slough, was a relatively narrow, broken stand of riparian forest with little of the grape or poison oak lianas so characteristic of the other sites. A comparison of the vegetation composition revealed a less diverse canopy and midstory. The canopy was composed almost exclusively of cottonwood. The relatively open midstory was dominated by Oregon ash.
Trapp (1972, 1978) documented ringtail anatomical and behavioral adaptations to climbing, as well as their ability to fully utilize all accessible aspects of the terrestrial environment. The ability to exploit the vertical aspect of their environment may allow ringtails to take advantage of a dense midstory, thereby increasing mobility about the forest. During times of winter flooding this could be especially important. A reduction, then, in a potentially critical aspect of their environment such as density of midstory vegetation might manifest itself in a smaller population size. This may partially explain variations in ringtail abundance.
Study sites II, IV, and V had similar ringtail densities and relatively diverse canopy and dense midstory vegetation layers. All three sites contained contiguous stands of riparian forest. Although the vegetation at study site I appeared to be physiognomically similar to that at study sites II, IV, and V, ringtail density was lower. This probably reflects a less intensive trapping effort (table 3).
There are few accounts in the literature on the abundance of ringtails and other procyonids. Grinnell etal . (1937) relied on the observations of W.H. Parkinson for density estimates. On 26 km2 (10 mi2 ) of land near Tollhouse, Fresno County, Parkinson trapped 31 ringtails within a season. Near the same area and over a 0.8-km (0.5-mi.) trail, Parkinson captured 13 ringtails. At 1,067 m. (3,500 ft.) on the western slope of Provo Mountain, near the Tuolumne River, within 0.65 km2 (0.25 mi2 ), he discovered five ringtail nests in blue oak trees. Each was occupied by a single ringtail. With information such as this, Grinnell etal . (ibid .) surmised that there was one ringtail per 13 km2 (5 mi2 ) throughout its range and in a few favored localities there were 2.3 per km2 (6 per mi2 ).
Taylor (1954) reported that J.D. Bankston, a trapper estimated ringtails at 3.9 per km2 (10 per mi2 ) in suitable broken country of the Edwards Plateau region in Texas.
Using home range data, collected with the use of radio-telemetry, Trapp (1978) estimated the density of ringtails in pinyon/juniper, blackbrush, and riparian vegetation of Zion Canyon, southwest Utah, to be 1.5–2.9 per km2 (3.8–7.6 per mi2 ).
Ringtail densities of 10.5–20.5 per 100 ha. (26.7–52.8 per mi2 ) documented during this study are the highest reported in the literature. Assuming that riparian woodland has higher productivity per unit area compared to chaparral and chaparral/woodland vegetations, the variation in ringtail density from shrub-dominated vegetationtypes to riparian woodland might be explained on the basis of this factor. A direct relationship
may also exist between ringtail density and physiognomic complexity (e.g., stratification) of the riparian plant community. Some evidence exists to support this contention from studies of the coati (Nasuanarica ), also a procyonid. The coati exploits arboreal aspects of its habitat (Davis 1960). Lanning (1976) reported densities of 1.2–2.0 coatis per 100 ha. in mixed evergreen woodland in and near Chiricahua National Monument, southeast Arizona. Densities of 42 and 26 coatis per 100 ha. have been estimated for tropical forests in Panama (Kaufmann 1962, cited by Lanning 1976). Lanning indicates that such differences in density may be attributed, in part, to ecological differences between the semiarid evergreen woodland and the moist tropical forests.
Acknowledgments
Special thanks are extended to Paul Laubacher, Randy Gray, Bob Motroni, Linda Heath, and particularly Dennis Messa for assistance in the field. Thanks to Gordon Gould, California Department of Fish and Game, for his cooperation and for arranging for the printing and mailing of the Furbearer Observation Questionnaire. Bob Motroni provided helpful suggestions of the techniques used for vegetational analysis. Larry Salata is gratefully acknowledged for critically reading and providing useful suggestions for this paper.
Literature Cited
Cottam, G., and J.T. Curtis. 1956. The use of distance measures in phytosociological sampling. Ecology 37:451–460.
Cox, G.W. 1976. Laboratory manual of general ecology. 232 p. Wm. C. Brown Co., Dubuque, Iowa.
Davis, W.B. 1960. The mammals of Texas. Texas Game and Fish Commission, Austin. Bulletin No. 41. 252 p.
Grinnell, J., J. Dixon, and J.M. Linsdale. 1937. Furbearing mammals of California. 2 vol., 777 p. University of California Press, Berkeley.
Hall, E.R. 1981. The mammals of North America. 2nd ed., 2 vol. 1181 p. John Wiley & Sons, New York, N.Y.
Hall, E.R., and K.R. Kelson. 1959. The mammals of North America. 2 vol., 1083 p. Ronald Press Co., New York, N.Y.
Ingles, L.G. 1965. Mammals of the Pacific states. 506 p. Stanford University Press, Stanford, Calif.
Kaufmann, J.H. 1962. Ecology and social behavior of the coati, Nasuanarica , on Barro Colorado. University of California Publ. Zool. 60:95–222 (cited by Lanning 1976).
Küchler, W.A. 1977. The map of the natural vegetation of California. p. 909. In : M.G. Barbour and J. Major (ed.). Terestrial vegetation of California. 1002 p. John Wiley & Sons, Inc. New York, N.Y.
Lanning, D.V. 1976. Density and movements of the coati in Arizona. J. Mamm. 57(3):609–611.
Long, C.A., and H.B. House. 1961. Bassariscusastutus in Wyoming. J. Mamm. 42(2):274–275.
Michny, F.J., D. Boos, and F. Wernette. 1975. Riparian habitats and avian densities along the Sacramento River. California Department of Fish and Game Administrative Report No. 75–1. 42 p.
Motroni, R.S. 1978. Sacramento Valley critical riparian habitat inventory. California Department of Fish and Game unpublished manuscript. 43 p.
Motroni, R.S. 1979. Avian density and composition of a riparian forest, Sacramento Valley, California. M.S. Thesis, California State University, Sacramento. 172 p.
Naylor, A.E., and G.W. Wilson. 1956. Unusual occurrence of the ring-tailed cat. Calif. Fish and Game 42(3):231.
Odum, E.P. 1971. Fundamentals of ecology. 3rd ed. 574 p. W.B. Saunders Co., Philadelphia, Penn.
Roberts, W.G., J.G. Howe, and J. Major. 1977. A survey of riparian forest flora and fauna. p. 3–19. In : A. Sands (ed.). Riparian forests in California: their ecology and conservation. 122 p. Institute of Ecology, University of California, Davis.
Schempf, P.F., and M. White. 1977. Status of six furbearer populations in the mountains of northern California. 52 p. USDA Forest Service Publications, California Region.
Seton, E.T. 1929. Lives of game animals. Vol. 2, Part I. Doubleday Doran & Co., Inc., Garden City, N.Y.
Smith, R.L. 1980. Ecology and field biology. 3rd ed. 835 p. Harper & Row Pub., New York, N.Y.
Stone T.B. 1976. Observations on furbearers within the riparian habitat of the upper Sacramento River. California Department of Fish and Game Memorandum Report. 12 p.
Taylor, W.P. 1954. Food habits and notes on life history of the ring-tailed cat in Texas. J. Mamm. 35(1):55–63.
Trapp, G.R. 1972. Some anatomical and behavioral adaptations of ringtails, Bassariscus astutus . J. Mamm. 53(3):549–557.
Trapp, G.R. 1978. Comparative behavioral ecology of the ringtail and gray fox in southwestern Utah. Carnivore 1(2):3–32.