7. Activity Patterns
Even without other knowledge, a description of the activity pattern of an animal reveals much about its life. As in human endeavors, the amount of time animals spend in various activities is a direct reflection of their basic economics. The activity of treeshrews was recorded by following radio-tagged individuals, rain or shine, from before they left the nest to begin their daily activity until they returned to the nest at its end. The results below are compiled from 185 complete daily records (see chapter appendix). Because it was rare to see the animals, the “activity” described here is an analysis of the movements of radio transmitters through the forest.
LENGTH OF THE ACTIVE PERIOD
All Ptilocercus individuals were active only at night, and all Tupaia individuals were active only by day. The time that activity began was almost fixed, but the time that it ended was highly variable.
Pentails left their nest high inside their giant hollow home tree and came down the tree after complete darkness had fallen, at a mean time of 1840 h (18.67 ± 0.31, N = 26; range: 18.27–19.17 h). There seemed to be a shift in exit time corresponding to the small shift in sunset during the year: activity began at a mean time of 1825 h in November and December (N = 7); at 1858 h from March through June (N = 9); and at 1833 h in August and September (N = 8). Because a different pentail was
All individuals of all Tupaia species left their nests just as the first gray light touched the forest floor, most at 0550 to 0555 h (the canopy lightens a few minutes earlier than does the ground). Exits were tightly clustered between 0545 and 0605 h. Adjustments to the length of the active period occurred through variation of times of return to the nest (fig. 7.1), never by changing the time that activity was initiated.
Pentails had the tightest grouping of times of return to the nest (fig. 7.1), with half of all records in the 15 minutes from 0515 to 0529 h and three-fourths of records within the last half hour of darkness, 0500 to 0530 h. When pentails returned to their sleeping site, the sky was already pale from the breaking dawn and there was far more light than there was when they emerged at nightfall, when to me it seemed pitch dark.
The diurnal Tupaia species were more variable in their behavior. Because emergence times were virtually constant, differences in the total length of the active period largely reflect differences in times of return to the nest. Females all had modal times of return to the nest within 15 minutes of 1800 h, but median times were earlier, reflecting the two- to threehour spread in time when animals retired. For most species, there are too few records to distinguish the behavior of males and females, but for T. tana, males usually entered the nest 1.25 hours earlier than females. (See table 7.1, fig. 7.1.) When total length of the daily active periods are compared within species, those of males are significantly different from those of females only for T. tana (Mann-Whitney U test P < 0.00). In betweenspecies comparisons, of females only, Ptilocercus is significantly different from all other species and T. tana from all but T. longipes (Mann-Whitney U test P < 0.05). These latter two are active for fewer hours per day than the others.
RESTING
During the day, Tupaia species rested from time to time. Those that I saw (T. tana and T. minor) crouched on a branch or log, with the tail curled snugly forward around the feet. They rested with eyes wide open, completely immobile and blending invisibly into the background. Lesser treeshrews that were in fruit trees would rest in a spot sheltered by leaves or branches from above and periodically get up, scratch and groom, eat a
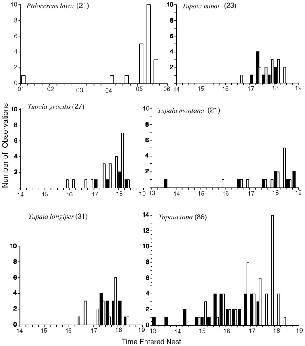
Fig. 7.1. Times at which radio-collared treeshrews entered their nests at the end of activity, grouped by 15 min intervals. Females = open bars; males = shaded bars.
Tupaia species were never known to rest by day in a nest, although a few times T. minor rested in a tree where there was a sleeping site and thus could have been in a nest (it was impossible to precisely pinpoint
| Species | Total Active Period(h) | SD | No. of Days | Time Resting(h) | Median No. of rests | No.of Days | End Activity Median(h)a |
|---|---|---|---|---|---|---|---|
| NOTE: Mean total length of active period (nest to nest); time recorded as resting (inactive for two consecutive triagulations or more) per active period (means of all complete tracking periods including those with no rests); median number of rests recorded per active period; and median time at which treeshrews returned to the nest at the end of activity. | |||||||
| *Beginning of 15-minute period into which median falls. | |||||||
| *Probability = 0.06; | |||||||
| **Probability = 0.04 (Mann-Whitney U). Males significantly different from females. | |||||||
| P. lowii, all | 10.29 | 1.09 | 20 | 1.61 | 1 | 20 | 0515 |
| T. minor, all | 11.81 | 0.56 | 20 | 1.25 | 1.5 | 20 | 1730 |
| Females | 11.88 | 0.61 | 13 | 1.23 | 2 | 13 | 1745 |
| Males | 11.68 | 0.47 | 7 | 1.29 | 1 | 7 | 1730 |
| T. gracilis, all | 11.70 | 0.65 | 24 | 0.71 | 1 | 24 | 1745 |
| Females | 11.64 | 0.65 | 19 | 0.86* | 1** | 19 | 1745 |
| Males | 11.89 | 0.67 | 5 | 0.13 | 0 | 5 | 1800 |
| T. longipes, all | 11.66 | 0.73 | 28 | 0.61 | 1 | 28 | 1730 |
| Females | 11.59 | 0.83 | 20 | 0.70 | 1 | 20 | 1715 |
| Males | 11.84 | 0.35 | 8 | 0.39 | 1 | 8 | 1745 |
| T. montana, all | 12.07 | 0.77 | 19 | 0.63 | 1 | 17 | 1800 |
| Females | 11.87 | 0.81 | 8 | 0.84 | 0.5 | 8 | 1800 |
| Males | 12.21 | 0.75 | 11 | 0.45 | 1 | 9 | 1800 |
| T. tana, all | 10.92 | 1.05 | 74 | 0.67 | 1 | 72 | 1645 |
| females | 11.15 | 0.94 | 53 | 0.70 | 1 | 54 | 1715 |
| Males | 10.34 | 1.11 | 21 | 0.57 | 1 | 18 | 1600 |
The two younger female pentails had a strikingly different behavior from that of any Tupaia: on nine of eighteen nights monitored they returned to the nest tree in the middle of the night for long rest periods. The adult female (F181) also did this on one of eight nights that she was followed. The young female F96 returned to the nest tree to rest, dividing her activity into two parts, on five nights between 22 November and 1 December, but on the last three nights she was tracked, between 8 and 30 December, she stayed away from the nest all night. This seems to show that forays out and back to the nest in pentails might be a juvenile behavior that largely disappears with independence. The other young female returned to the nest for midnight rests on four nights, interspersed between five nights during which she did not return to the nest at all.
The distribution of longer rests, detectable by the radio-tracking method I used, is the inverse reflection of activity (fig. 7.2). All species concentrated resting toward the middle of the active period, but only pentails showed a sharp, unimodal, peak of mid-activity rests. These long rests divided the night into two well-defined peaks of activity during the first and last thirds. All the Tupaia species were 100 percent active only in the first two to four hours of the morning, and they would rest sporadically almost any time after that. All species have a bimodal tendency for inactivity, with rests most often in late morning and midafternoon. The inverse, overall activity is basically bimodal when viewed cumulatively, especially in T. longipes, for which resting was most strongly clustered; but within the midday period there were two minor activity peaks.
These averaged trends do not show what an individual treeshrew does in a day, which is better reflected in the statistics (fig. 7.2, table 7.1). While tracking Tupaia on foot from daybreak to dark, matching their movements on a parallel trail, it immediately becomes obvious that they seldom rest. The two statistics, median number of rests per day and mean total time resting recorded per day, together show that the four largest species rest, on average, less than an hour each day and most often only once. That is, on many days no rests at all were recorded. The amount of time spent resting was not significantly different between any species pairs, but the difference between the sexes was significant for T. gracilis, for number of rests, and nearly so for time spent resting (with more data it might reach significance). Short rests of less than 20 minutes may have
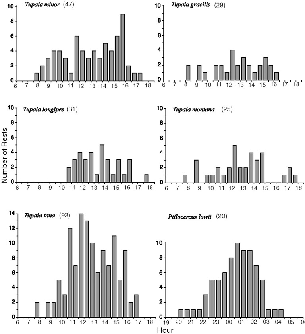
Fig. 7.2. Distribution of inactivity recorded during radio-tracking. Excludes rainstorms. Rest period scored in the 30 min period during which its major part occurred; if an inactive period was longer than 30 min, it is scored into each half hour of which it formed a major part. Y axis = number of times a treeshrew rested in that period. N = number of inactive periods recorded for the species, not the number of 30 min periods in which resting was recorded.
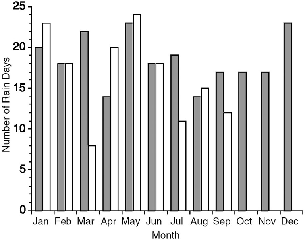
Fig. 7.3. Number of days per month on which it rained at Danum Valley in 1990 (shaded bars) and 1991 (open bars).
RAIN
In the equable temperatures of lowland equatorial forests, rain is the main climatic event that influences animal activity. The pentails foraged near their nest tree during continuously rainy nights and returned to it if nearby at the start of heavy downpours. Tupaia species ceased activity during heavy rains, usually after a short movement to a nearby spot, evidently a known shelter. If a storm struck in the afternoon, treeshrews often went to a nest and stayed there for the remainder of the day. Rarely, a tupai made straight for a nest and retired when black sky, thunder, and winds presaged an afternoon storm. If it was raining when treeshrews were due to emerge at dawn (or dusk for pentails), they sometimes delayed the start of activity, but eventually they started to move around in moments of lighter rain.
Because rain caused treeshrews to cease activity, I took advantage of the detailed rainfall records from the Research Centre hydrology project
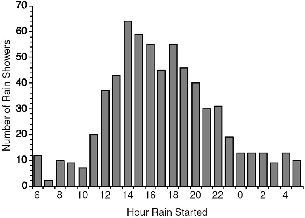
Fig. 7.4. Hour of onset of rain at Danum Valley, all recorded showers.
At Danum Valley Field Centre in 1990, it rained on 222 days, or almost two-thirds of days, and in 1991 it rained on 149 days from January to September (fig. 7.3; see fig. 3.5 for monthly rainfall). In 1990 rain fell for more than 339.77 hours (this is the sum of available records, but data on length of a few rains were lost because of chart-recorder problems or failure to change the paper, although data for the amount of rainfall are complete). Per month, it rained for from 10.92 hours (April) to 56.58 hours (December), or 7.9 percent of the time.
The degree to which rain might truncate an animal's activity depends on which part of the day it occurs. The hours during which most rains began (fig. 7.4) and the hours in which it fell (fig. 7.5) varied somewhat with the season. Throughout the year it mainly rained in the afternoon or first hours of the night. Only in December did rains also start in the last half of the night and continue into the morning. From July through October rains were bimodal, usually starting either near midday or early afternoon or just after nightfall (fig 7.4). In June alone, rain onset was concentrated only in the evenings between 16 h and 20 h. For Tupaia species, December was the month during which maximal rain (10.67 h)
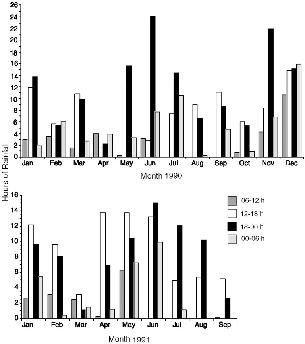
Fig. 7.5. Number of hours during which rain fell during each quarter of the day, for each month of the study. Some records are missing from the data because of chart recorder problems.
Pentails are faced with a high probability of rain at the hour when they become active (figs. 7.4, 7.5). In June and November 1990, 22 to 24 hours of rain occurred during the first half of the night (18–00 h), or up to 13 percent of those hours, and in January, May, July, and December more than 13 hours of rain fell between 18 h and 00 h. At the maximum in June, it rained for a least 32.43 hours during the active periods of pentails. When I monitored pentails during torrential evening rains (from a tarp shelter next to the nest tree), I found that they at first remained inside the tree but were active within it and then emerged during the rain for short forays into neighboring large trees, where perhaps they were able to forage within sheltered nooks or beneath giant branches.
DISCUSSION OF ACTIVITY PATTERNS
The outstanding feature of treeshrew activity is the sheer amount of it. With known rests subtracted, the least active species (P. lowii) spent a mean of 8.7 hours each day in motion, while the most active species (T. montana) averaged 11.44 hours in motion. However, short rests that were not detected and rests enforced by rain decreased these totals by unquantified amounts. Treeshrews spent little time with others, and as corroborated by direct sightings, most of their activity seemed to be concerned with foraging or feeding, especially in the case of females (see chaps. 8, 9). For most Tupaia species, the modal time of entry to the nest was within an hour of nightfall, but the large variation of return times (fig. 7.1) implies that when treeshrews had eaten enough, they retired for the rest of the day. This could be field tested experimentally, by artificial provisioning.
The largest species, T. tana, has a shorter period of activity than the others, and the second largest, T. longipes, tends to a lesser extent to retire earlier than yet smaller species (table 7.1). This likely reflects the smaller relative metabolic needs associated with larger body size; but in that case food acquisition time would likewise have to be scaled, as absolute needs would be greater in larger species. The much longer activity of T. tana females than males is I believe associated with reproductive
Pentails have a relatively shorter total period of activity than do the diurnal species of the same small body size. Since most pentails finally retired only at dawn, their short activity was due both to long midnight rests and to the delay in exit from the nest tree until pitch darkness, half an hour or more after nightfall. The ability to maintain themselves with shorter activity than small Tupaia may be the result of a low metabolic rate and torpor while inactive (Whittow and Gould 1976). In captivity West Malaysian pentails kept by Gould (1978) likewise became active quite late (1945–2015 h), suggesting that this pattern is intrinsic to pentail behavior. By rising late, pentails could avoid exposure to crepuscular predators such as owls, which hunt most effectively in dim light.
In a study of T. minor in West Malaysia (observations from a fixed site), D'Souza (1972) found that lesser treeshrews began activity very late (0745 h), two hours after the 0545 h sunrise, and then exhibited a sharply bimodal activity pattern with no activity from 12 to 14 h, a peak in late afternoon, and return to the nest at about 1800 h. This contrasts with my data from Sabah, which show exit from the nest universally at dawn and resting taking place any time from late morning to late afternoon, with a slight trend of trimodality in activity. D'Souza saw evidence that after days of heavy rain, lesser treeshrews began activity earlier, presumably to compensate for time lost, and he speculated that late exit from the nest represented slack time that could be used if needed. If lesser treeshrews have shorter active periods and longer rests in West Malaysia than on Borneo, the former site may have higher food availability and reduced obligate foraging time. Unfortunately, the differences in our methods preclude direct comparison. In Sabah it was clear that if treeshrews had “extra” time, they cut activity short at the end of the day, never at the beginning.
The daily need for food must be greatest when treeshrews emerge from their nests following their twelve-hour sleeping phase. For pentails, emergence from the nest in almost all months coincides with the time of day with most likelihood of rainstorms (fig. 3.5). Frequent rains at nightfall could restrict foraging at the most critical time of the night and hence could reduce the ability to acquire extra nutrients for reproduction. Their low metabolic rate may help pentails cope with foraging delays or deprivation caused by rain. Because of their timing, rains at Danum seemed
The direct effects of rain on animals have rarely been studied, and detailed rainfall records are available for few sites. For tropical rainforest birds, it has been suggested that the breeding seasons exclude the rainiest months because heavy rains both physically destroy nests and curtail vital foraging time (Brosset 1990, and pers. com.; Foster 1974). If birds breed during the wet season, rain may reduce the clutch size that parents can feed (Foster 1974).
In summary, the daily activity cycle of Tupaia species in Sabah, as monitored by radio-tracking, shows sustained activity and relatively little rest during the day. This implies that foraging takes most of the day. Ptilocercus lowii had both a shorter active phase and more resting time than the Tupaia but nonetheless foraged nightly for about nine hours. These behaviors imply that the energetic balance of treeshrews may be tight, a hypothesis for which I give more evidence in the next two chapters.
EVOLUTIONARY NOTES
Long daily foraging times in treeshrews are linked to their short, simple digestive tracts and rapid passage times (chap. 5). Food must be ingested repeatedly, in short bouts throughout the daily active phase. In captivity at the National Zoo, T. tana from Sabah became obese when fed ad libitum and developed dewlaps and aprons of fat (Miles Roberts pers. com.; pers. obs.), whereas in the wild no body fat was ever noted. In the same captive circumstances, T. minor did not become obese. Captive common shrews (Sorex araneus) also put on much body fat, but wild shrews are never fat (Churchfield 1990). That wild treeshrews were always lean and foraged (moved actively about) for nine to eleven hours of each twelve-hour day suggests that they need long daily foraging times just to break even energetically. Tupaia species sometimes retired to their nests before the end of the daylight period, so rather than more foraging, they opted for shelter, away from the risk of predation by raptors and perhaps other predators. As predicted from theory concerning body size and metabolic rate, the larger the treeshrew species, the more likely it was to enter the nest early.
The smallest nonflying insectivores, the true shrews, are destined by their tiny body sizes and high relative metabolic rates to frequent search for food. Churchfield reported,
Most shrews must feed regularly every two or three hours or they will die of starvation, so they must remain active both day and night. Periods of activity are spent bustling through the undergrowth or along subterranean tunnels as they search for prey and explore their home ranges. Bouts of activity may last from about 30 minutes to two hours, and alternate with periods of rest in the nest. (1990: 123)
Common shrews have about ten bouts of activity per twenty-fourhour day (Churchfield 1990). Treeshrews seem to have a similar, high activity pattern (although with few, or short, rests), but it is restricted to half of the day. Most small mammals, including treeshrews, most tenrecs (Eisenberg and Gould 1970), elephant shrews (Rathbun 1979), marsupials (Flannery 1990; pers. obs.), prosimians (Doyle and Martin 1979), bats, and most but not all rodents (volelike grassland herbivores are among the exceptions) have evolved nocturnal or diurnal lifestyles with twelve-hour activity cycles. This permits these species to tune (by evolution) their physiology and sensory systems to optimal function for their activity phase and to become more efficient at feeding on resources with different day or night characteristics. Cathemeral (active day and night) activity in mammals appears to be found only in species that are unable to acquire enough nutrition within one of the twelve-hour phases, from constraints of either digestive/acquisition systems (grazers), or search time for rare or hard to catch prey (small felids), or metabolic constraints (shrews). Other benefits of twelve-hour activity include shelter from predation for at least half of the twenty-four-hour day and whatever physiological advantages long sleep confers.
APPENDIX: METHODS
We triangulated the location of a treeshrew every ten to twenty minutes, or more often if it was moving rapidly. The number of locations logged per day thus depended both on how many hours the animal was active and on how quickly and far it moved; but most daily samples were of about 40 points. For most of the above analyses, data are included only from the 185 entire days that were not grossly curtailed by heavy rains.
The beginning of activity, when an animal left its nest, was usually precisely signaled by a clear change in intensity or pattern of the radio signal. I often listened continuously to capture this moment. The instant of return to the nest was sometimes also readily identified (as when an animal went underground or climbed to a high nest), but at other times it had to be inferred from cessation of activity for the rest of the day. If a treeshrew went to its nest early in the day, I waited in the field and monitored it until the end of the day, to make sure that it did not resume activity. The beginning of activity was determined on a greater
During tracking, we often recorded our perception of whether an animal was active or inactive, and for transmitters with activity monitoring options, the transmitter activity signal was noted. However, for the analyses in this chapter, I do not use these measures but instead define inactivity, or “rest,” as any two sequential triangulations between which the animal did not move. This standardizes the treatment of the data and allows more of it to be used, but obviously rests of shorter than twenty minutes were not detected, so that the time spent resting is underestimated. To economize batteries, we turned off the radio receivers between triangulations. For treeshrews with activity sensors, it would have been better to sample some days by running the receiver continuously to measure all inactive periods exactly.