3. The Milieu:
Field Study Sites
and Habitats
One of two Malaysian states on Borneo, Sabah is a quiet, rural region that occupies the island's northern tip, sandwiched between the South China and Sulu seas (fig. 3.1). Borneo was once totally forested, except above the treeline on mountains, but much of its surface is now agricultural, silvicultural, or heavily logged. The sparse human population of Sabah has put relatively little pressure on the land, and the large tracts of forest that remain hold some of the last, best populations of Sumatran rhinos, proboscis monkeys, and orangutans. Beside these well-publicized species lives a rich fauna of little-known smaller mammals (Payne, Francis, and Phillipps 1985). The biota of Borneo reflects its history as part of Sundaland, when during Pleistocene glaciations it was joined by dry land to the continent of Southeast Asia, when sea levels were 160 m lower than at present. Its fauna thus includes species that are the same as, or closely allied with, those of Sumatra, peninsular Malaysia, and Thailand. The biological diversity of Borneo is much enhanced by high mountain ranges with habitat gradients from lowlands to cloud forests, including Mount Kinabalu, the highest mountain in Southeast Asia. About 20 percent of the mammal species of Borneo are endemic to it, including seven of its ten species of treeshrews. All of these treeshrews dwell in the tropical rainforests, and the large geographic extent and elevational variation of Borneo may have provided a field for speciation and persistence of this elsewhere poorly represented family.
Fig. 3.1. Location of study sites.
Like tropical forests everywhere, the appearance and structure of the forests of Sabah vary considerably from place to place, depending on soil type, slope, drainage, rainfall, exposure, history of disturbance, and biogeographic history. Nevertheless, on well-drained stable ridge tops and slopes grows a forest that is one of the greatest wonders of the biological world. Taller, and with more large trees than other tropical rainforests, the Asian dipterocarp forests are in stature akin to the giant Sequoia forests of the United States; but whereas only two or three tree species form the Sequoia canopy, more than three hundred species may crowd a single hectare in Borneo (Newbery et al. 1992; Primack and Hall 1992). Many emergents (trees that emerge above the canopy) reach 50 m in height; a few tower to above 75 m (fig. 3.2). These giants are mostly of the dipterocarp family (Dipterocarpacae) and a legume (Koompassia excelsa). However, where soil conditions are unfavorable to the dipterocarp giants, a Bornean forest looks much like tropical rainforests on other continents worldwide: it may be tall and open below; irregular in height and undergrowth; or short and sparse in canopy and snarled with festoons of lianas or thickets of vicious rattan climbers. Treeshrews live in all forest types on Borneo.

Fig. 3.2. An immense, canopy-size dipterocarp tree on a logging road near Poring. Note Alim Biun and Hajinin Hussain standing in the road to the left of the base of the tree.
PORING HOTSPRINGS RANGER STATION
The first phase of field studies was from March to August 1989, at Poring Hotsprings Ranger Station (Poring), in Kinabalu Park, Sabah, Malaysia (06°03$$N; 116°42$$E; fig. 3.1), under the hospitality of the Sabah Parks Department. Two of the park staff, Alim Biun and Hajinin Hussain, collaborated full time in the research during this field season, to the great benefit of the study. Alim and Hajinin cheerfully endured barbaric work schedules (sometimes radio-tracking from 4:30 in the morning to 8:30 at night) and taught me much about the animals and forest. At Poring we worked on two study plots, a “low” plot on a ridge crest at the base of the Eastern Ridge of the Mount Kinabalu massif and a “high” plot on a ridge 2.5 km northwest from Poring, above the trail to Langanan Falls.
The low plot occupies a small side ridge separated by a steep ravine and stream from the main Eastern Ridge. We chose this site because it included the largest relatively flat area of lowland forest in that section of the park. All vegetation below it was secondary or agricultural. The plot began at about 535 m elevation at the lower limit of the mature forest and extended 500 m in a wedge to where the ridge narrowed to about 50 m at 720 m elevation. We worked along a trail above this 300 m farther, to 800 m elevation. On this area we opened and used a small trail system for radio-tracking, which was flanked by steep ravines on each side, and we trapped treeshrews in a grid on the wider, lower part (fig. 3.3). The flattish triangle within our trails measured 6 ha and was 200 m wide at the base and 460 m long. We followed treeshrews in this plot throughout our study at Poring.
This lowland plot had vegetation characterized by a high canopy of emergents, dominated by tall dipterocarps on the higher well-drained ridge tops and edges, with generally smaller trees such as oaks (Lithocarpus spp., Castanopsis spp.) occupying the lower portions. The understory ranged from bare and open to vine-choked ravines and thickets of saplings in old lightgaps (openings caused by treefalls). Most of the study plot had a fairly open understory. A high diversity of dense rattan palms scrambled throughout much of the forest, and groves of understory Licuala sp. and other palms grew at 700 m on slopes (fig. 3.4). Where light entered on ravine sides or rocky slopes, curtains of vines draped the steep rock faces. Except in dank ravine bottoms, there were scarcely any trunk or branch epiphytes on the trees at this elevation, apart from isolated baskets of giant staghorn ferns (Platycerium sp.). The forest floor had a good cover of leaf litter, except where this was swept away
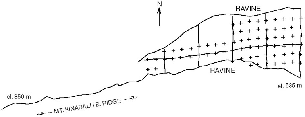
Fig. 3.3. Map of trails and trap sites (crosses) on the lower plot at Poring.
The Langanan (high) plot was likewise chosen for its flattish profile and workable vegetation, after initial trapping showed it to include many montane treeshrews. It also was on a ridge top, in the shape of a rough equilateral triangle encompassing 3.8 ha. It ranged in elevation from 930 to 1,000 m. We filled the area with lines of traps spaced at 25 m intervals. We worked in this plot only to study mountain treeshrews (T. montana), from 29 June to 31 July 1989. To reach the high plot to start work before dawn, we had to leave our house at 0430 h and walk for an hour.
The forest on the high plot is a closed canopy of small- to mediumdiameter trees, generally small-leafed, growing on a rocky substrate (fig. 3.5). The understory is dense and characterized by numerous, prehensile rattans that grab one's skin and clothing and enforce cautious movement. At this elevation condensation from the afternoon mists, which daily shroud Mount Kinabalu during many months of the year, is evident in an abruptly increased density of trunk and branch epiphytes such as moss, ferns, and orchids; moss on the rocks and tree trunks; and the formation of a spongy mat of tree roots and moss lying above the soil surface. At the level of the plot this springy surface root mat was just beginning to develop and quite thin. At higher elevations it can become several feet thick, with the moss-covered root mat entirely above ground

Fig. 3.4. The forest understory at 600 m on the study plot at Poring.
The lands skirting Mount Kinabalu are quite densely occupied by Dusun people, for whom hunting is an important tradition. Poaching in the park was intense and difficult to control, and large game mammals were much reduced or in some cases extirpated in the vicinity of Poring, so that the fauna there appeared to be diminished at the time of our study. Much of the forest outside of the park has been cut, leaving few natural areas for legal hunting, and there were no significant tracts of lowland forest workable from Poring.
DANUM VALLEY
In 1990 I moved the project to Danum Valley Conservation Area (Danum), in eastern Sabah near Lahad Datu (4°58'N, 117°48'E; fig. 3.1), chiefly because the steep, ravined terrain at Poring made it very difficult to track the weak transmitters carried by small mammals. It also seemed preferable to work in a site with less hunting and a more intact and tranquil fauna. I studied treeshrews in the field at Danum Valley Field Centre from 29 August 1990 to 2 October 1991. At Danum an able botanist,

Fig. 3.5. The forest at about 900 m on the high study plot at Poring, habitat of T. montana.

Fig. 3.6. Helicopter view of Danum Valley Field Centre. The treeshrew study plot is in the forest above the river in the right half of the photo. The slender suspension bridge can just be seen.
Danum Valley Field Centre and its 43,800 ha reserve of pristine forest, which comprises the Danum Valley Conservation Area, has been the base for extensive field research. This has allowed my studies on treeshrews to be embedded into a far more complete ecological background than could have been possible elsewhere in Sabah. Some of these studies are collected in a recent publication (Marshall and Swaine 1992) that includes outlines of the history of the field station and its forests, soils, and climate Marsh and Greer 1992. Although poaching has severely
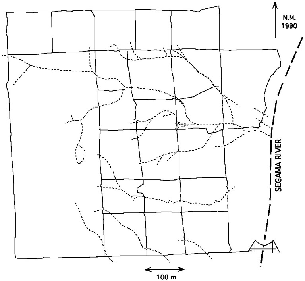
Fig. 3.7. Map of the trails (solid lines) and streams (dotted lines) on the study plot at Danum Valley.
The study plot at Danum is within the conservation area adjacent to the Segama River, just north of the bridge, and ranges from about 450 m elevation at the river to 650 m elevation at its top (northwest) corner. It occupies the east-facing slope of the west (left) bank of the river basin and includes two permanent and several temporary small tributary streams. The terrain is a rolling series of erosion gullies and ridges perpendicular to the river on the flank of a long, gentle ridge that reaches 900 m elevation about 2 km west of the river. I chose this as the focal study area because it had already been the base plot for studies of the bird fauna (Lambert 1992) and litterfall and litter fauna (Burghouts et
The vegetation of the study area is classified as lowland evergreen dipterocarp forest Marsh and Greer 1992. It has a high tree species diversity, with 511 species (>Io cm DBH = 2,248 individuals) identified in a 4 ha parcel next to my study plot, of which the 2 ha halves had 388 and 387 species, respectively (Newbery et al. 1992). Dipterocarpaceae was the second best represented tree family in the parcel, with 9.3 percent of individuals (about 100/ha); but Euphorbiaceae strongly dominated, with 27.5 percent of all trees (about 300/ha), and it would more appropriately be called a euphorb than a dipterocarp forest. My adjacent study plot has large disturbed areas and also seemed to have fewer large trees than the Newbery parcels, perhaps because of the effects of wind and erosion (treefalls) on the river-facing slopes and because the Newbery plots are on a quite flat, well-drained ridge top.
The treeshrew study area is a mosaic of tall open forests on welldrained ridgelines, intersected with moist, densely vegetated gullies and a small ravine. The canopy generally has a high layer of emergents (40–60 m tall) and large trees (30–40 m) that are separated by wide gaps through which light penetrates to fall on a closed subcanopy formed by the tops of small trees and lianas many meters below (fig. 3.8). The true, connected closed canopy is often very low, at 6 to 10 m, although there is large variation in its height. This structure, with lack of closure of the high canopy, may be created or favored by the hilly terrain, which precludes formation of a uniform canopy surface and allows some side penetrance of light. The understory on the plot includes almost no rattans or other palms and varies between open on well-drained ridge tops to totally closed in old treefall gaps. There are few epiphytes, and the forest floor has in most places a well-developed standing layer of leaf litter (production about 3.4 t/ha/yr;Burghouts et al. 1992. The open river and its valley exposes the basin to winds that are desiccating during dry months. Four large areas of about a third to half a hectare each have no tree canopy and are smothered with nearly impenetrable tangles of old lianas. The largest such area covers a rocky substrate much like a boulder

Fig. 3.8. Helicopter view of the forest canopy on the study plot at Danum Valley. Note how tall, isolated trees emerge above a lower, more closed tree canopy below.
For comparison with results from mature forest at Danum, I made a brief trapping survey in logged forest outside the conservation area 1 km northeast of the field center. This forest had been selectively logged from primary forest in 1989. It was a mosaic of relatively little damaged areas that still had some tree canopy and extremely disturbed sections, open to the sky, with many vine blankets and stands of Macaranga and Vismia.
RAINFALL AND FRUITING PHENOLOGY
There were no climate or rainfall records for Poring Hotsprings Ranger Station at the time of my study, but detailed climate records have been kept since 1985 by the stream hydrology project at Danum Valley (see Marsh and Greer 1992). Mean annual rainfall there from 1986 to 1990 was 2,822 mm. Monthly rainfall shows no strongly predictable pattern,
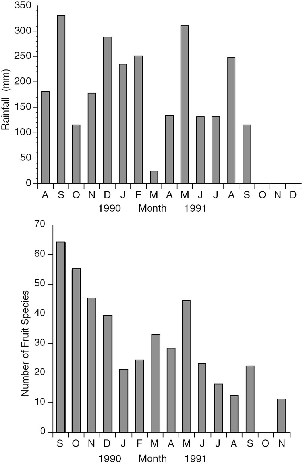
Fig. 3.9. Monthly rainfall during the study period at Danum Valley (top) (courtesy of the Danum Hydrology Project); number of fruit species collected per month on study plot trails (bottom).
To evaluate the seasonal patterns in a treeshrew resource at Danum Valley, Gasis worked with me to collect and identify all fallen fruit from a transect that included the grid of 4.5 km of trails encompassing most of the home ranges of the radio-collared study animals. The complete list of fruit species that we identified is compiled in Appendix I.
The overall fruiting phenology showed two peaks of fruit abundance during the calendar year (fig. 3.9), centered in September and March, the rainiest months during the study. By good fortune, the start of our work at Danum coincided with the first major fruit-masting episode recorded at the Field Centre since its inception in 1984, so we were able to document the consequences of a rare fruiting event. Unfortunately we missed the beginning of the masting phenomenon and only monitored its decline from the peak period (September-December 1990; fig. 3.9). More fruit species were collected in each of the four months during the masting period than in any of the ten following months. The number of species of fallen fruit recorded during masting in September 1990 (63) was three times that of September 1991 (22), and November 1990 had four times as many fruit species as the same month in 1991.
APPENDIX: METHODS
Most methods germane to specific topics are detailed in appendixes in the relevant chapters, and only a few general techniques are given here. The statistical analyses and some graphs were done with Systat© 5.2 for the Macintosh, and most graphs were drawn with DeltaGraph© Pro 3.
Fruit Sampling Methods. We collected fallen fruit during two days near the beginning of each month from September 1990 through November 1991, except October 1991. All fruit in each 100 m trail section was collected together into a separate plastic bag, so that information on the number of fruit species, and total weight of fruit, could be evaluated in terms of distance. Our fruit survey is but a subsample of what was available on the study area. Although there are a number of obvious biases created by the methodology itself, the method should give a good reflection of relative monthly values (fig. 3.9B).
The outstanding feature of the rainfall records for the year before the masting phenomenon was an exceptionally wet May and June, with 600 to 700 mm of rain falling in each, almost double the usual amount, following a very dry April (about 30 mm). These rains perhaps triggered the mass flowering that subsequently produced massive fruiting.
In August and September 1991 dense, acrid smoke, evidently from burning rice fields in Kalimantan hundreds of kilometers away, blanketed Danum and most of Sabah. Airports were temporarily closed, and visibility was reduced to a couple of hundred meters. It is possible that blockage of the light could have influenced plant phenology, or rains falling through smoke may have acidified the streams, but no data are available.